營養學/脂類的生化代謝
| 醫學電子書 >> 《臨床營養學》 >> 脂類 >> 脂類的生化代謝 |
| 臨床營養學 |
|
|
|
唾液中無消化脂肪的酶,胃液中雖含有少量的脂肪酶,但成人胃液酸度很強,不適於脂肪酶的作用,故脂肪在成人口腔和胃中不能消化。嬰兒胃液的PH在5左右,奶中脂肪已經乳化,故脂肪在嬰兒胃中可消化一部分。脂肪的消化主要在小腸內進行。食糜通過胃腸粘膜產生的胃腸激素刺激胰液和膽汁的分泌,並進入小腸。膽汁中的膽鹽是強有力的乳化劑,脂肪受到膽鹽的乳化,分散為細小的脂肪微粒,有利於和胰液中的脂肪酶充分接觸。
胰液中的胰脂肪酶能將部分脂肪完全水解為甘油和游離脂肪酸,但有一半的脂肪僅能局部水解為甘油二酯或甘油一酯。因為胰脂肪酶能特異地和連續地作用於甘油三酯的1和3位置,開始解脫一個脂肪酸,形成甘油二酯。然後,再解脫一個脂肪酸,形成甘油一酯。胰脂肪酶對甘油三酯的水解率和其脂肪酸鏈的長短有關,不飽和脂肪酸比飽和脂肪酸易於水解。還有小部分的脂肪完全不水解。
脂肪的水解產物游離脂肪酸和甘油一酯、甘油二酯進入腸粘膜細胞內,在滑面內質網上重新合成與體內脂肪組成成分相近的甘油三酯。新合成的甘油三酯的組成和構型適宜於以後的代謝。新合成的甘油三酯在粗內質網上與磷脂、膽固醇、蛋白質形成乳糜微粒,經腸絨毛的中央乳糜管匯合入淋巴管,通過淋巴系統進入血液循環。水解產物甘油因水溶性大,不需膽鹽即可通過小腸粘膜經門靜脈而吸收入血液。完全未被水解的脂肪亦能以乳膠微粒的形式直接進入腸粘膜細胞,在內質網上合成的乳糜微粒再由淋巴系統進入血液循環。因此,動物和植物脂肪幾乎完全吸收。食後2h,可吸收24~41%,4h後吸收53~71%,6h後吸收68~86%,12h後吸收97~99%。
影響脂肪吸收的因素:
(1)脂肪的熔點脂肪的熔點會影響其吸收。例如,羊脂的熔點為44~55℃,其吸收率為85%;而椰子油的熔點為28~33℃,其吸收率為98%。一般說來,植物油的熔點較低,所以較易吸收。這是因為進入十二指腸中的脂肪應該是液態,這樣才能乳化。脂肪的熔點比體溫越高,就越難於乳化,所以也就越不容易消化吸收。
(2)脂肪攝取量因為脂肪吸收比較慢,小量食入時吸收率高。大量時有不少排泄掉,吸收率低。
(3)年齡1歲內的嬰兒脂肪吸收率低,常易發生消化不良。老年人脂肪的吸收和代謝都比年輕人慢。
(4)脂肪酸組成一般來說,含短鏈脂肪酸的脂肪其吸收比長鏈的為快。含奇數碳鏈脂肪酸的脂肪,其吸收比偶數的為慢。棕櫚酸在甘油第2位的脂肪(如豬油、人乳),其吸收比棕櫚酸在其他位置或在第2位的其他脂肪酸的脂肪(牛油、羊油、牛乳)來得好。
(5)鈣脂肪吸收時,雖然需要一定量的鈣,但如鈣量過高時,則脂肪吸收反而下降。特別是含月桂酸、豆蔻酸、軟脂酸和硬脂酸等熔點高的脂肪。而含油酸和亞酸這些不飽和脂肪酸濃度高的脂肪,鈣不影響其他吸收。
鈣干擾飽和脂肪吸收的機理是由於形成難溶解的飽和脂肪酸鈣。
3.3.2 磷脂的消化和吸收
卵磷脂在小腸內由四種酶進行分解,由胰腺分泌的磷脂酶A原,受胰蛋白酶激活成磷脂酶A,在膽鹽和Ca2+存在下,作用於卵磷脂,釋出一個脂肪酸,產生溶血卵磷脂。它有溶血作用。
磷脂酶B作用於卵磷脂,釋出二分子脂肪酸,產生α-甘油磷酸膽鹼。溶血卵磷脂亦可受磷脂酶B的作用,釋出一分子脂肪酸後,生成α-甘油磷酸膽鹼。
最後,甘油磷酸酶和膽鹼磷酸酶分別作用於α-甘油磷酸膽鹼,完全水解成甘油、磷酸及膽鹼。
除脂肪酸外,磷脂的消化產物大多數是水溶性的,在腸道內易於吸收。
小部分磷脂在膽鹽的協助下,混合在乳膠微粒內,在腸內可以不經消化而能直接吸收。但大部分磷脂仍需水解後才被吸收。吸收的磷脂水解產物,也可以在腸壁重新合成完整的磷脂分子再進入血液中。
3.3.3 膽固醇的消化和吸收
食物中所含的膽固醇,一部分是與脂肪酸結合的膽固醇酯,另一部分是游離狀態的。胰液和腸液中均含有膽固醇酯酶,在腸道內催化膽固醇脂水解,產生游離的膽固醇和脂肪酸。
膽固醇為脂溶性物質,故必須藉助膽鹽的乳化才能在腸內吸收。但是吸收的膽固醇約有三分在腸粘膜細胞內經酶的催化重新酯化,形成適合體內需要的膽固醇酯。再與部分未酯化的游離膽固醇、磷脂、甘油三酯及由腸粘膜細胞合成的脫輔基蛋白一起形成乳糜微粒,經淋巴系統進入血液循環。因此,淋巴和血液中的膽固醇大部分以膽醇酯的形式存在。
影響膽固醇吸收的因素:
(1)食物膽固醇在攝食1、3或6g膽固醇後,其吸收率分別為60、40、30%,即吸收率隨著攝食量的增加而遞減。這是因為增加膽醇吸收的同時,發生兩個代償機制,即膽固醇排泄增加和體內合成減少。
(2)食物脂肪和脂肪酸食物中的脂肪和脂肪酸具有提高膽固醇吸收的作用。這是由於:①乳糜微粒中的膽固醇主要是膽固醇酯,脂肪和脂肪酸可以在腸粘膜中供給膽固醇以再酯化所需要的脂醯基,從而有利於膽固醇吸收;②高脂肪膳食具有促進膽汁分泌的作用。而膽汁中的膽汁鹽能促使膽固醇形成微粒,有助於膽固醇的吸收。
(3)植物固醇各種植物固醇,如豆固醇、谷固醇等,不僅其本身吸收很差,而且還能抑制膽固醇的吸收。有人認為這可能是因為:①植物固醇的分子結構與膽固醇極為相似,競爭性抑制腸內膽固醇酯的水解,以及腸壁內游離膽固醇的再酯化,促使其由糞便中排泄;②植物固醇競爭性地佔據微粒內膽固醇的位置,影響膽固醇與腸粘膜細胞接觸的機會,從而妨礙其吸收。
(4)其它食物中不能被利用的多糖,如纖維素、果膠、瓊脂等容易和膽汁鹽結合形成複合物,妨礙微粒的形成,故能降低膽固醇的吸收。此外,腸細菌能使膽固醇還原為不易吸收的糞固醇。因此,長期服用廣譜抗生素的病人,常能增加膽固醇的吸收。
3.3.4 脂肪的合成
脂肪合成有兩條途徑:一是利用食物中的脂肪轉化而成人體脂肪;另一是將糖轉變為脂肪,這是體內脂肪的主要來源。脂肪組織和肝臟是體內脂肪合成的主要場所。合成脂肪的原料是磷酸甘油和脂肪酸。
磷酸甘油是由糖代謝的中間產物磷酸丙糖還原而成,或從食物中消化吸收的甘油在甘油激酶的催化下,與ATP作用而生成磷酸甘油。
脂肪酸用於脂肪合成之前需經活化,即在脂肪醯CoA合成酶的催化下,與輔酶A、ATP作用生成脂醯輔酶A。二分子脂醯輔酶A經過脂醯轉移酶的催化。將脂醯基轉移到α-磷酸甘油分子上,生成α-磷酸甘油二酯,又稱磷脂酸。後者經磷脂酸酶作用,脫去磷酸後再與另一分子脂醯輔A在甘油二酯轉脂醯酶的作用下,結果就生成脂肪。
3.3.5 磷脂的合成
體內磷脂一部分是直接由食物中來,另一部分是在各組織細胞內,經過一系列酶的催化而合成。磷脂的種類很多,現僅將甘油磷脂和神經磷脂的合成簡述如下:
(1)甘油磷脂的合成甘油磷脂主要是在微粒體內合成,其原料為磷酸、甘油、脂肪酸、膽鹼或乙醇胺等。其中必需脂肪酸由食物供應,其他原料可在體內合成。蛋白質分解產生的甘、絲及蛋氨酸即可作原料。甘氨酸在體內經亞甲基四氫葉酸作用變為絲氨酸,再脫羧變乙醇胺,由S-腺苷蛋氨酸供給甲基而變為膽鹼。膽鹼和ATP在膽鹼磷酸激酶催化下生成磷酸膽鹼,再和胞苷三磷酸(CTP)在胞苷酸轉移酶作用下變為胞苷二磷酸膽鹼(CDP-膽鹼),他和甘油二酯在磷酸膽鹼轉移酶催化下脫掉胞苷一磷酸(CMP),就形成α-卵磷脂。
腦磷脂的合成與卵磷脂的合成過程基本相似,不同的的只是以乙醇胺代替膽鹼。
磷脂還可以從另一條途徑合成,即α-磷酸甘油二酯先與CTP作用生成CDP-甘油二酯,再與絲氨酸反應生成磷脂醯絲氨酸,後者直接脫羧即生成腦磷脂。腦磷脂甲基化即可生成卵磷脂。
(2)神經磷脂的合成人體內的神經磷脂種類很多,現僅將腦醯胺為中間產物的一種合成過程列於圖3-4。
3.3.6 膽固醇的代謝
(1)膽固醇的合成人體內的膽固醇一部分(約40%)是由動物性食物中來,稱為外源膽固醇;一部分是由體內組織細胞自行合成,稱為內源膽固醇。
人體內幾乎所有組織都具有合成膽固醇的能力,但合成的速率和總合成量並不一致,肝臟不僅合成快,而且合成量也最多,是人體合成膽固醇最活躍的場所,其次是小腸。
合成的原料可來自乙醯輔酶A。前後經30多步酶促反應,全部過程在細胞質內進行。
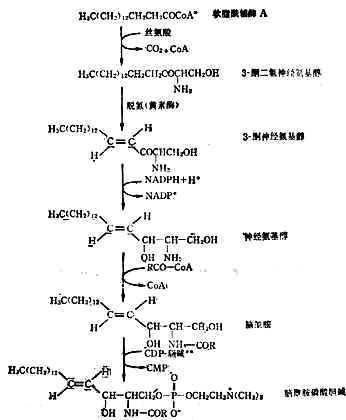
圖3-4 腦醯胺磷酸膽鹼的合成途徑
為了儲存和轉運的目的,游離膽固醇和長鏈脂肪酸結合成膽固醇酯。組織中的膽固醇是在膽固醇醯基轉移酶的作用下,接受脂醯CoA的脂醯基形成膽固醇脂。血漿膽固醇的酯化是在血漿中的卵磷脂膽固醇基轉移酶(LCAT)催化下進行的。
(2)膽固醇合成的調節在調節膽固醇的合成中,β-羧-β-甲戊二酸(HMG)-CoA還原酶具有決定性意義,任何因素改變還原酶的活性時,則顯著影響體內膽固醇的合成。食入高膽固醇後,當膽固醇含量升高時,可反饋抑制肝臟膽固醇的合成。此作用主要是還原酶活性下降之故。這種負反饋機制只發生於大鼠和其它動物的肝臟,而在胃腸道卻沒有發現。至於人體,外源性膽固醇不能降低肝外組織的膽固醇合成。其對肝臟的影響,仍有不同的意見。因此,大量攝食膽固醇後,血漿膽固醇仍有一定程度升高。
膽固醇合成速率在晝夜之間可相差4~5倍,午夜時合成最快,上午10時左右最慢,這與還原酶活性的變化是一致的。
還有許多激素也對此酶有影響,如腎上腺素和去甲腎上腺素能促進此酶的合成,因此使膽固醇合成增加。甲狀腺素能增加組織對兒茶酚胺的敏感性,而後者又增加HMG-CoA還原酶的合成。但甲腺素又能增加膽固醇的分解和排出,而後者的作用卻大於前者,結果使血清膽固醇降低。故甲亢病人常見血漿膽固醇降低,而甲狀腺機能減退病人則血漿膽固醇增高。cAMP也有抑制膽固醇合成的作用,於是有人推測有些激素的作用是通過cAMP的變化而實現的。
肝臟內膽固醇的合成還受脂肪代謝的影響。當脂肪酸動員加強,不僅血甘油三酯升高,膽固醇合成也明顯增強。運動能使血漿游離脂肪酸含量減少,從而使膽固醇合成緩慢。多不飽和脂肪酸可在體內轉變成前列腺素,有人認為前列腺素可通過cAMP-蛋白質激酶系統而使HMG-CoA還原酶失活,或通過抑制脂肪動員,降低血漿游離脂肪酸而使肝中膽固醇合成減少。這可能是多不飽和脂肪酸降低血漿膽固醇的一個重要原因。
糖、脂肪和蛋白質分解而產生的乙醯CoA主要經三羧酸循環徹底氧化,但過量的乙醯CoA不僅可以合成脂肪,也可在微粒體酶系的作用下大量生成HMG-CoA。因此,HMG-CoA還原酶的酶促反應隨之加快,膽固醇合成增加。
(3)膽固醇的轉化膽固醇在體內能轉變成一系列有生理活性的重要類固醇化合物:①轉變為膽汁酸。膽固醇在肝臟內受7α-羥化酶的催化生成7α-羥膽固醇,後者經一系列反應轉變為膽汁酸。這是膽固醇的主要去路,人體內約有80%的膽固醇可以在肝臟中轉變為膽汁酸,其中主要是膽酸、脫氧膽酸和鵝脫氧膽酸。膽酸再與甘氨酸或牛磺酸結合成膽汁酸。膽汁酸以鈉鹽或鉀鹽的形式存在,稱之為膽鹽。它們對脂類的消化吸收起重要作用;②轉變為維生素D3。在肝臟和腸粘膜細胞內,膽固醇可轉變為7-脫氫膽固醇。後者經血液循環運至皮膚,再經紫外光照射,7-脫氫膽固醇可轉變為維生素D3。維生素D3能促進鈣磷吸收,有利於骨胳生成;③轉變成類固醇激素。膽固醇在腎上腺皮質細胞內轉變成腎上腺皮質激素,如醛固酮,皮質醇。再卵巢內可轉變為孕酮與雌激素。在睾丸內可轉變為睾酮。
(4)膽固醇的排泄部分膽固醇可以在組織內(主要是肝臟)還原,生成二氫膽固醇,與膽固醇一起分布於全身各組織。體內膽固醇由肝臟排入膽汁,隨膽汁進入腸腔。一部分通過腸肝循環重新吸收入肝臟。一部分在腸道被經細菌作用後轉變為糞固醇。未被吸收的二氫膽固醇以及很難吸收的的糞固醇統稱為中性糞固醇,隨糞便排出體外。
此外,尚有少量膽固醇和二氫膽固醇以皮脂形式由皮膚排出;小部分膽固醇和膽固醇脂隨表皮細胞脫落,一起排出體外。
3.3.7 脂蛋白的代謝
血漿脂蛋白包括乳糜微粒、極低密度脂蛋白、低密度脂蛋白和高密度脂蛋白。它們的代謝簡述如下:
(1)乳糜微粒(CM)的代謝CM的90%是甘油三酯,其餘為磷脂、蛋白質和膽固醇。食物脂肪的水解產物經小腸吸收,並在小腸粘膜上皮細胞滑面內質網中重新合成甘油三酯。這些甘油三酯和從食物吸收的磷脂、膽固醇結合,同時與糙面內質網合成的脫輔基蛋白(apo)B和A-I形成原始的CM。它經過高爾基體加工後進入淋巴系統,稱為淋巴CM。淋巴CM到達血液後,在組成成分和結構方面都發生一些改變。它主要包括接受由HDL轉移來的apoC,同時還向血漿中釋放磷脂和吸收膽固醇,這種CM遂變為成熟CM。
當血液經過脂肪組織、肝臟、肌肉等的毛細血管時,經管壁脂蛋白脂酶的作用,可使CM中的甘油三酯水解成脂肪酸和甘油。這些水解產物的大部分則進入細胞被利用或重新合成脂肪而儲存。由於CM失去中心部分的甘油三酯而逐漸變小,這種CM稱為CM殘餘。此時CM表面上的脫輔基蛋白、磷脂、膽固醇都脫離而移到HDL上。此種作用進行得很快,所以正常人空腹血漿幾乎不易檢出CM。有人認為CM殘餘可能在肝中轉變為LDL。
(2)極低密度脂蛋白(VLDL)的代謝VLDL主要由肝實質細胞合成,其合成及分泌過程與小腸粘膜上皮細胞合成CM的過程基本相似。VLDL主要成分也是甘油三酯,但磷脂和膽固醇的含量比CM的多。其蛋白質部分除apoB以外,還有apoCⅠ、Ⅱ、Ⅲ,apoE,少量的apoAⅠ、Ⅱ和apoD。
肝細胞合成VLDL的甘油三酯,其來源是由糖在肝細胞中轉變而來,也可由脂庫中脂肪動員出來的游離脂肪酸在肝細胞的滑面內質網中重新合成。所以,VLDL是轉運內源性脂肪的主要運輸形式。此外,糙面內質網合成apoB,再與質膜的磷脂形成複合體,最後在高爾基體內結合成VLDL。它所含的膽固醇酯的來源不十分明了,可能是HDL輸送來的。
VLDL和CM一樣,經肝外脂蛋白脂酶的作用,使其中的甘油三酯水解成脂肪酸和甘油,被細胞利用或重新合成甘油三酯而儲存。由於甘油三酯減少,膽固醇相對地增多,VLDL的結構遭受破壞。ApoC脫離這種已經改變的脂蛋白,使脂蛋白脂酶的作用停止。與此同時,LCAT酶被apoA-Ⅰ和apoC-Ⅰ激活,開始發揮催化作用,將一部分卵磷脂的β-脂醯基轉移至膽固醇分子上,前者成為溶血卵磷脂,後者成為膽固醇酯。溶血卵磷脂的極性較大,便釋放於血漿中。膽固醇沒有極性,於是轉移到脂蛋白的核心部分。經過上述變化,脂蛋白的體積比原來縮小,其所含的總膽固醇相對地增多,甘油三酯和apoC相對減少,但仍能維持一定的球形,通常把這種VLDL殘骸稱為中間低密度脂蛋白(ILDL)。
ILDL的成分是:apo15%,膽固醇29%~33%(其中75%為酯型),磷脂17%,甘油三酯35~39%。與VLDL比較,它的酯型膽固醇含量較高,而甘油三酯含量較低。
(3)低密度脂蛋白(LDL)的代謝VDLD水解產物ILDL到達肝臟,與肝細胞膜上的ILDL受體結合,ILDL遭受分解,釋放出「多精肽」與其它脂類,最後變成LDL。人體的VLDL不是全部的話,亦是絕大部分轉變為LDL。
與VLDL相比,LDL的膽固醇酯增多,apoB幾乎沒有減少,甘油三酯顯著下降,而除apoB以外的蛋白質、磷脂等成分都除去了。所增加的膽固醇酯,可能是由於HDL的膽固醇經LCAT的作用而酯化,並轉移到LDL上來的。
LDL的分解主要在肝外實質細胞中進行。細胞表面上有LDL的特異的受體,LDL通過它的apoB的正電荷精氨酸殘基與此受體結合。結合的LDL在細胞表面的凹陷結構內為表面膜包成小泡,被攝入細胞內。含LDL的小泡與細胞內的溶酶體融合,LDL的蛋白質被溶酶體內的蛋白酶水解為胺基酸。酸性脂肪酶將膽固醇酯分解為游離膽固醇和脂肪酸。這種游離膽固醇能抑制能微粒體內HMG-CoA還原酶的活性,因而阻止細胞內膽固醇的合成。同時,又使微粒體內的LCAT酶活化,使膽固醇轉變為適於儲存的膽固醇酯(圖3-5)。當細胞內游離膽固醇水平增高以及膽固醇酯開始堆積時,LDL受體的合成受到抑制,LDL吸收率下降。這樣能維持血漿中和細胞內膽固醇濃度的平衡。
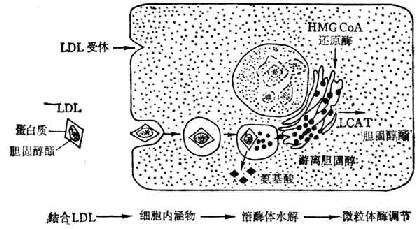
圖3-5 人體成纖維細胞內LDL降解步驟
在正常人體內每天降解的LDL為總量的45%,其中約有三分是通過LDL受體途徑進行的。此外,還有另一個降解途徑,就是通過清除細胞(Scavenger cells)的吸入和降解LDL。這種細胞是從事於非特異性的胞飲作用。由這條途徑所降解的血漿LDL是個常數(15%)。
(4)高密度脂蛋白(HDL)的代謝在肝臟或小腸內,CM經脂蛋白脂酶的作用,將甘油三酯分解,水解產物及其表層的磷脂、游離膽固醇和apoA離開CM而形成雙層脂類組成的顆粒,這就是新生HDL,它呈碟形。這種新生HDL進入血液後,其表面被來自周圍組織和其它脂蛋白的大量游離膽固醇所佔據。此時,這些顆粒成為LCTA的最適底物,在LCTA催化下,使表面的游離膽固醇轉變為膽固醇酯,並由顆粒表面轉入顆粒核內。因核內脂類含量劇增,內壓升高,使磷脂雙層壓變為單層。原有的碟形排列消失,形成一種富含膽固醇酯的成熟球形顆粒。這樣,富含游離膽固醇的碟形新生HDL就轉變為富含膽固醇酯的球形成熟HDL。
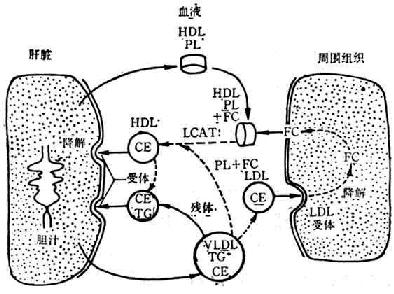
FC-游離膽固醇 CE-膽固醇酯 PL-磷脂 TG-甘油三酯
圖3-6 HDL代謝特點
由於HDL代謝的特點,它能從周圍組織轉運膽固醇到肝臟進行降解排泄(圖3-6)。這樣,能防止膽固醇沉積在血管壁上,甚至已經沉積的膽固醇,亦能由HDL予以轉移,可以防止並有可能消除動脈粥樣硬化的形成。
肝臟和小腸是HDL的主要降解部位。血液中的成熟HDL,和細胞膜上的受體結合,進入細胞內,由溶酶體予以降解。分解出來的膽固醇一部分不變,一部分轉變為膽酸而從膽汁排出。少量的HDL亦可在腎臟、腎上腺、卵巢等器官內降解。
| 關於「營養學/脂類的生化代謝」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |