生理學/腎上腺皮質
| 醫學電子書 >> 《生理學》 >> 內分泌 >> 腎上腺 >> 腎上腺皮質 |
| 生理學 |
|
|
腎上腺皮質分泌的皮質激素分為三類,即鹽皮質激素、糖皮質激素和性激素。各類皮質激素是由腎上腺皮質不同層上皮細胞所分泌的,球狀帶細胞分泌鹽皮質激素,主要是醛固醇(aldosterone);束狀帶細胞分泌糖皮質激素,主要是皮質醇(cortisol);網狀帶細胞主要分泌性激素,如脫氫雄酮(dehydroepiandrosterone)和雌二醇(estradiol)也能分泌少量的糖皮質激素。腎上腺皮質激素屬於類固醇(甾體)激素,其基本結構為環戊烷多氫菲。鹽皮質激素與糖皮質激素是21個碳原子的類固醇,雄激素含有19個碳原子,雌激素含有18個碳原子(圖11-15)。
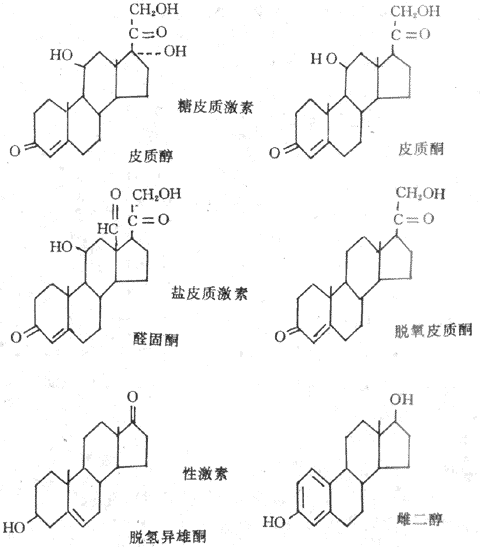
圖11-15 幾種主要的腎上腺皮質激素有化學結構
膽固醇是合成腎上腺皮質激素的原料,主要來自血液。在皮質細胞的粒線體內膜或內質網中所含的裂解酶與羥化酶等酶系的作用下,使膽固醇先變成孕烯酮,然後再進一步轉變為各種皮質激素。由於腎上腺皮製裁各層細胞存在的酶系不同,所以合成皮質激素亦不相同(圖11-16)。
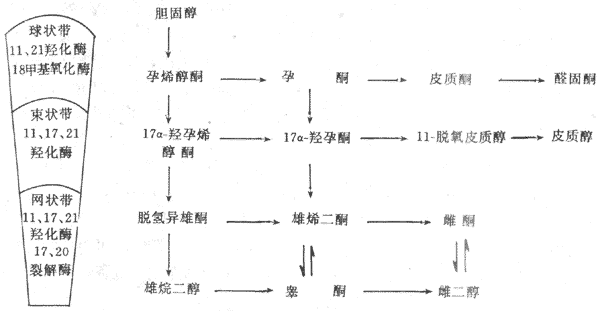
圖11-16 腎上腺皮質激素合成的主要步驟
皮質醇進入血液後,75%-80%與血中皮質類固醇結合球蛋白(corticosteroid-binding globulin,CBG)或稱為皮質激素運載蛋白結合,15%與血漿白蛋白結合,5%-10%的皮質醇是游離的。結合型與游離型皮質醇可以相互轉化,維持動態平衡。游離的皮質醇能進入靶細胞發揮其作用。CBG是肝產生的α2球蛋白,分子量為52000,血漿中CBG濃度為30-50mg/L。CBG與皮質醇有較強有親和力,每一分子的CBG僅有一個結合位點,只能結合一個分子的皮質醇。每100ml血漿CBG能結合20μg皮質醇。可見,CBG在運載皮質醇方面起著重要作用,。醛固醇與血漿白蛋白及CBG的結合能力很弱,主要以游離狀態存在和運輸。
皮質醇在血漿中半衰其為70min,醛固醇為20min。它們都在肝中降解,皮質醇首先是在C4GN C5間的雙鍵加氫還原,形成雙氫皮質醇,隨後,C3上的酮基變成羥基產生四氫皮質醇,與葡萄糖醛酸或硫酸結合,隨尿排出體外。四氫皮質醇是皮質醇的主要代謝產物,點尿中皮質醇代謝物排出量的45%-50%。四氫皮質醇和皮五醇在C20酮基變為痙基生成皮五醇,占尿中排出量的20%左右。由於四氫皮質醇和皮五醇在C17上均有羥基,故稱為17-羥類固醇。另外,C1`7上脫去側鏈,生成17-氧類固醇,占尿中排出題的10%左右。醛固醇基本上循類似途徑被處理。
腎上腺皮質網狀帶分泌的性激素以脫氫異雄酮為主,它是一種17-氧類固醇,睾酮的代謝產物也是17-氧類固醇。因此,男子尿中17-氧類固醇的來源有睾丸分泌的睾酮和腎上腺皮質分泌的皮質醇及雄激素。
(二)腎上腺皮質激素的生物學作用
動物摘除雙側腎上腺後,如不適當處理,一二周即死去,如僅切除腎上腺髓質,動物可以存活較長時間,說明腎上腺皮質是維持生命所必需的。分析動物死亡的原因,主要有兩個方面:其一是機體水鹽損失嚴重,導致血壓降低,終於因循環衰竭而死,這主要是缺乏鹽皮質激素所致;其二是糖、蛋白質、脂肪等物質代謝發生嚴重紊亂,對各種有害刺激的抵抗力降低,導致功能活動失常,這是由於缺乏糖皮質激素的緣故。若及時補充腎上腺皮質激素,動物的生命可以維持。
1.糖皮質激素 人體血漿中糖皮質激素主要為皮質醇,其次為皮質酮,但皮質酮的含量僅為皮質醇的1/20-1/10。
(1)對物質代謝的影響:糖皮質激素對糖、蛋白質和脂肪代謝均有作用。①糖代謝;糖皮質激素是調節機體糖代謝的重要激素之一,它促進糖異生,升高血糖,這是由於它促進蛋白質分解,有較多的胺基酸進入肝,同時增強肝內與糖異生有關酶的活性,致使糖異生過程大大加強。此外,糖皮質激素又有抗胰島素作用,促進血糖升主。如果糖皮質激素分泌過多(或服用此類激素藥物過多)可引起血糖升高,甚至出現糖尿;相反,腎上腺皮質功能低下患者(如阿多數狄森病),則可出現低血糖;②蛋白質代謝:糖皮質激素促進肝外組織,特別是肌肉組織蛋白質分解,加速胺基酸轉移至肝生成肝糖原。糖皮質激素分泌過多時,由於蛋白質分解增強,合成減少,將出現肌肉消瘦、骨質疏鬆、皮膚變薄、淋巴組織萎縮等;③脂肪代謝:糖皮質激素促進脂肪分解,增強脂肪酸在肝內氧化過程,有利於糖異生作用。腎上腺皮質功能亢進時,糖皮質激素對身體不同部位的脂肪作用不同,四肢脂肪組織分解增強,而腹、面、肩及背有脂肪合成有所增加,以致呈現面圓、背厚、軀幹部發胖而四肢消瘦的特殊體形。
(2)對水鹽代謝的影響:皮質醇有較弱的貯鈉排鉀作用,即對腎遠由小管及集合管重吸收和排出鉀有輕微的促進作用。此外,皮質醇還可以降低腎小球進球血管阻力,增加腎小球血漿流量而使腎小球濾過率增加,有利於水的排出。皮質醇對水負荷時水的快速排出有一定的作用,腎上腺皮質功能不足患者,排水能力明顯降低,嚴重時可出現「水中毒」,如補充適量的糖皮質激素即可得到緩解,而補充鹽皮質激素則無效。有資料指出,在缺乏皮質醇時,ADH釋放增多,集合管對水的重吸收增加。
(3)對血細胞的影響:糖皮質激素可使血中紅細胞、血小板和中性粒細胞的數量增加,而使淋巴細胞和嗜酸性粒細胞減少,其原因各有不同。紅細胞和血小板的增加,是由於骨髓造血功能增強;中性粒細胞的增加,可能是由於附著在小血管壁邊緣的中性粒細胞進入血液循環增多所致;至於淋巴細胞減少,可能是糖皮質激素使淋巴細胞DNA合成過程減弱,抑制胸腺與淋巴組織的細胞分裂。此外,糖皮質激素還能促進淋巴細胞與渚酸性粒細胞破壞。
(4)對循環系統的影響:糖皮質激素對維持正常血壓是必需的,這是由於:①糖皮質激素能增強血管平滑肌對兒茶酚胺的敏感性(允許作用),這可能由於糖皮質激素能嗇血管平滑肌細胞膜上的兒茶酚胺受體數量以及調節受體介導的細胞內的信息傳遞過程;②糖皮質激素能抑制具有血管舒張作用的前列腺素的合成;③糖皮質激素能降低毛細血管的通透性,有利於維持血容量。腎上腺皮質功能低下時,血管平滑肌對兒茶酚胺的反應性降低,毛細血管擴張,通透性增加,血壓下降,補充皮質醇後可恢復。
另外,離體實驗證明,糖皮質激素可增強心肌的收縮力,但在整體條件下對心臟的作用並不明顯。
(5)在應激反應中的作用:當機體受到各種有害刺激,如缺氧、創傷、手術、飢餓、疼痛、寒冷以及精神緊張和焦慮不安等。血中ACTH濃度立即增加,糖皮質激素也相應增多。能引起ACTH與糖皮質激素分泌增加的各種刺激稱為應激刺激,而產生的反應稱為應激(stress)。在這一反應中,除垂體-腎上腺皮質系統參加外,交感-腎上腺髓質系統也參加,所以,在應激反應中,血中兒茶酚胺含量也相應增加。切珍重腎上腺髓質的動物,可以抵抗應激而不產生嚴重後果,而當去掉腎上腺皮質時,則機體應激反應減弱,對有害刺激的抵抗力大大降低,嚴重時可危及生命。應激反應可能從以下幾個方面調節機體的適應能力:
①減少應激刺激引起的一些物質(緩激肽、蛋白水解酶及前列腺素等)的產生量及其不良作用;②使能量代放運轉以糖代謝為中心,保持葡萄糖對重要器官(如腦和心)的供應;③在維持血壓方面起允許作用,增強兒茶酚胺對血管的調節作用。應該指出,在應激反應中,除了ACTH、糖皮質激素與兒茶酚胺的分泌增加外,β-內啡肽、生長素、催乳素、抗利尿激素、胰主血壓素及醛固醇等均可增加,說明應激反應是多種激素參與並使機體抵抗力增強的非特異性反應。
糖皮質激素的作用廣泛而複雜,以上僅簡述了它們的主要作用。此外,還有多方面的作用,如促進胎兒肺表面活性物質的合成,增強骨骼肌的收縮力,提高胃腺細胞對迷走神經與胃泌素的反應性,增加胃酸與胃蛋白酶原的分泌,抑制骨的形成而促進其分解等。臨床上使用大劑量的糖皮質激素及其類似物,可用於抗炎、抗過敏、抗毒和抗休克。
2.鹽皮質激素主要為醛固醇,對水鹽代謝的作用最強,其次為脫氧皮質醇(表11-3)。
表11-3 幾種腎上腺皮質激素對糖代謝作用的比較
| 激素 | 對糖代謝作用 | 保鈉排鉀作用 |
| 皮質醇 | 1.0 | 1.0 |
| 可地松 | 0.8 | 0.8 |
| 皮質酮 | 0.5 | 1.5 |
| 醛固酮 | 0.25 | 500 |
| 脫氧皮質酮 | 0.01 | 30 |
表中數字代表皮質激素的相對效力,以皮質醇的效力為1.0,即醛固酮的保鈉排鉀作用為皮質醇的500倍
醛固酮是調節機體水鹽代謝的重要激素,它促進腎遠曲小管及集合管重吸收鈉、水和排出鉀,即保鈉、保水和排鉀作用。當醛固酮分泌過多時,將使鈉和水貯留,引起高血鈉、高血壓和血鉀降低。相反,醛固酮缺乏時則鈉與水的排出過多,血鈉減少,血壓降低,而尿鉀排出減少,血鉀升高。關於醛固酮對腎的作用及其機制,可參閱第八章,另外,鹽皮質激素與糖皮質激素一樣,以增強血管平滑肌對兒茶酚胺的敏感性,且作用比糖皮質激素更強。
(三)腎上原皮質激素分泌的調節
1.糖皮質激素分泌的調節腎上腺皮質分泌皮質激素的束狀帶及網狀帶,處於腺垂體保腎上腺皮質激素(adrenocortiotropin,ACTH)的經常性控制之下,無論是糖皮質激素的基礎分泌,還是在應激狀態下的分泌,都受ACTH的調控,切除動物的垂體後,束狀帶與網狀帶萎縮,糖皮質激素有分泌顯著減少,如及時補充ACTH,可使已發生萎縮的束狀帶與網狀帶基本恢復,糖皮質激素有分泌回升。
(1)ACTH:ACTH是一個含39個胺基酸的多肽,分子量為4500,其化學結構示於圖11-17。
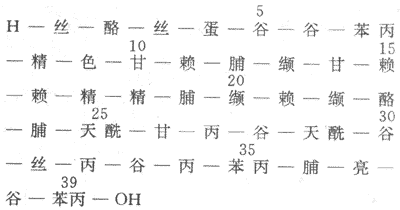
圖11-17 人ACTH的化學結構
ACTH分子上的1-24位胺基酸為生物活性所必需的,25-39位胺基酸可保護激素,減慢降解,延長作用時間。各種動物的ACTH前24位胺基酸均相同,因此,從動物(牛、羊、豬等)腺垂體提到的ACTH對人有效。目前,ACTH已能人工合成。在垂體,ACTH是由阿黑皮素原(POMC)經酶分散而來,同時產生β-MSH。ACTH再經酶分解生成α-MSH,ACTH的第4-10位胺基酸與α-MSH第4-10位胺基酸和β-MSH第11-17位胺基酸相同,這部分胺基酸是產生MSH活性最小單位,因此ACTH也具有促黑素細胞產生黑色素的作用。
ACTH的分泌呈現日節律波動,入睡後ACTH分泌逐漸減少,午夜最低,隨後又逐漸增多,至覺醒起床前進入分泌高峰,白天維持在較低水平,入睡時再減少。由於ACTH分泌的日節律波動,促糖皮質激素的分泌也出現相應的波動。ACTH分泌的這種日節律波動,是由下丘腦CRH節律性釋放所決定的。
ACTH 不但刺激糖皮質激素的分泌,也刺激束狀帶與網狀帶細胞的生長發育,關於ACTH的作用機制已基本清楚。在束狀還與網狀帶細胞膜上存在ACTH特異性受體,在Ca2+存在的條件下,ACTH與膜受體結合,激活腺苷酸環化酶,通過cAMP激活蛋白激酶,蛋白激酶起三項重要作用;①使核糖蛋白磷酸化,促進mRNA形成一種特殊蛋白質,使膽固醇得以進入粒線體,並經側鏈解形成孕烯醇酮,以進一步合成糖皮質激素;②使磷酸化酶活化,促進糖原分解,產生ATP,提供能量,另外還通過戊糖旁路產生還原型輔酶Ⅱ(NADPH),以利膽固醇的羥化過程;③使膽固醇酯活化,促進其轉變為膽固醇,提供激素合成的原料。在ACTH促進腎上腺皮質細胞合成糖皮質激素的同時,束狀帶細胞膜對葡萄糖與膽固醇的轉運機制增強,使較多的葡萄糖與膽固醇進入細胞內(圖11-18)。
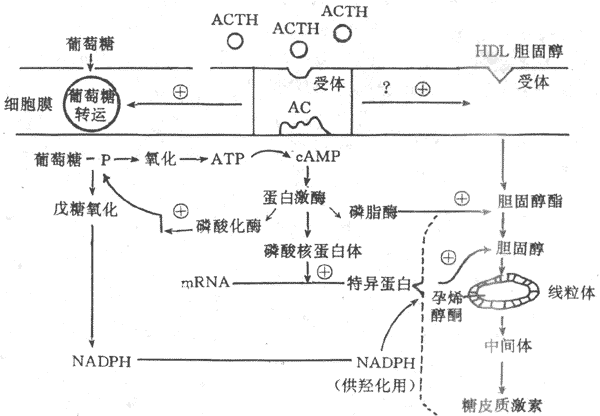
圖11-18 ACTH作用機制示意圖
HDL:高密度脂蛋白 AC:腺苷酸環化酶
(2)ACTH分泌的調節:ACTH調節糖皮質激素的分泌,而ACTH的分泌受下丘腦CRH的控制又與糖皮質激素有反饋調節。下丘腦CRH神經元和其他下丘腦調節肽神經元一樣,又受腦內神經遞質的調控。應激刺激作用於神經系統的不同部位,最後通過神經遞質,將信息彙集於CRH神經元,然後借CRH控制腺垂體的促腎上腺皮質激素細胞分泌ACTH。此外,當血中糖皮質激素濃度升高時,可使腺垂體釋放ACTH減少,ACTH的合成也受到抑制,腺垂體對CRH的反應也性減弱。糖皮質激素的負反饋調節主要作用於垂體,也可作用於下丘腦,這後一種反饋稱為長反饋。ACTH還可反饋抑制CRH神經元,稱為短反饋。至於是否存在CRH對CRH神經地的超短反饋,尚不能肯定。
綜上所述,下丘腦、垂體和腎上腺皮質組成一個密切聯繫、協調統一的功能活動軸,從而維持血中糖皮質激素濃度的相對穩定和在不同狀態下的適應性變化(圖11-19)。
2.鹽皮質激素分泌的調節 醛固酮的分泌主要受腎素-血管緊張素系統的調節。另外,血K+,血Na+濃度可以直接作用於球狀帶,影響醛固酮的分泌(詳見第四章與第八章)。
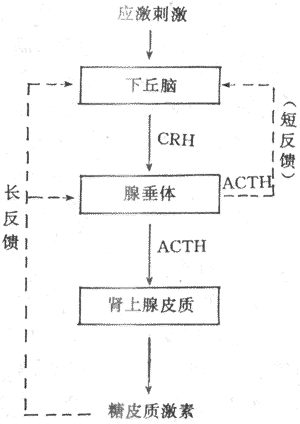
圖11-19 糖皮質激素分泌的調節示意圖
實線表示促進 點線表示抑制
在正常情況下,ACTH對醛固酮的分泌並無調節作用,但切除垂體後,在應激醛固酮的分泌反應減弱,提示在應激情況下,ACTH對醛固酮的分泌可能起到一定的支持作用。
參看
| 關於「生理學/腎上腺皮質」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |