營養學/碳水化物的生化代謝
| 醫學電子書 >> 《臨床營養學》 >> 碳水化物 >> 碳水化物的生化代謝 |
| 臨床營養學 |
|
|
|
4.2.1 消化吸收
碳水化物要消化或單糖才吸收。消化的過程就是水解的過程。麥芽糖、乳糖、蔗糖、麥芽低聚糖都能消化。人能消化的多糖僅澱粉一種,糖原在製成食品時已不存在了。
消化從口腔開始,口腔里有唾液澱粉酶能水解交替α1→4糖甘鍵,但不能水解α1→6糖苷鍵和相鄰的α1→6糖苷鍵。消化產物是糊精、麥芽低聚糖和麥芽糖。
胃裡沒有消化澱粉的酶。唾液澱粉酶的最適PH是6.6~6.8,在食糜沒有被胃酸中和以前,能持續作用一段時間,使澱粉和低聚糖能再消化一部分。
小腸內有胰液的α-澱粉酶,其作用和唾液澱粉酶相同,把直鏈澱粉消化成麥芽糖和麥芽三糖,支鏈澱粉消化成麥芽糖、麥芽三糖及由4~9個葡萄糖分子組成的而有α1→6苷鍵的麥芽低聚糖。
腸粘膜上皮細胞中有吸收細胞,每一細胞約有3000條微絨毛,微絨毛間的空間的有效半徑約0.4nm。只有上述消化產物能夠通過,與微絨毛膜上的酶反應。膜上的酶有四種:①α1→4糖苷酶,把葡萄糖分子自上述產物一個個地切下來;②異麥芽糖酶,水解麥芽低聚糖的α1→6糖苷鍵;③蔗糖酶,消化蔗糖;④β-半糖苷酶,消化乳糖。
所以消化分兩步進行:①腸腔內的消化,產物是雙糖和麥芽低聚糖;②微絨毛膜上的消化,產物是單糖。
微絨毛雙脂質層的(詳後)。四種酶嵌在雙脂質層內,活性位伸在膜外。在其近處,還有全部嵌在膜內的運輸單糖的蛋白質,這樣,消化的最終產物立刻可以被運輸蛋白所結合。
運輸蛋白在結合葡萄糖以前,先結合腸腔內的Na+排入腸腔,腸腔中Na+都帶入細胞內,釋放到胞漿中。Na+排入腸腔,腸腔中的Na+濃度比細胞內高,自低濃度排到高濃度要消耗能量,所需能量由ATP供應。糖進入細胞後,約有15%流迴腸腔,25%擴散入血,60%與靠近基膜一端的質膜上的另一載體蛋結合而離開細胞。這一結合不需Na+,而且運輸葡萄糖的速度比葡萄糖從腸腔進入吸收細胞的速度快,所以葡萄糖不會在吸收細胞中蓄積,從而提高了吸收效率。當食糜到達空腸下部時,95%的碳水化物都被吸收了。
4.2.2 中間代謝概述
小腸吸收的碳水化物主要是葡萄糖、果糖、半乳糖三種,經門靜脈送到肝臟。葡萄糖進入肝細胞後與磷酸反應生成葡萄糖-6-磷酸,這樣細胞內的葡萄糖濃度可維持在低水平,使葡萄糖不斷進入肝細胞。在吸收的葡萄糖中,60%以上在肝內代謝,其餘入大循環。果糖和半乳糖在肝中轉變為葡萄糖。
葡萄糖在肝內經分解代謝提供機體所需要的能量,多餘的合成糖原保留在肝內,再有多餘就轉變成脂肪運送到脂肪組織儲存起來。
葡萄糖的分解代謝分三個階段:
(1)酵解葡萄糖分解的第一階段是生成丙酮酸。這一系列反應和碳水化物在體外經酵母的作用生成丙酮酸相同,所以稱為酵解。所不同的是發酵時丙酮酸進一步生成灑精;在體內,有氧時丙酮酸氧化成二氧化碳和水,無氧時還原成乳酸。
酵解分步進行(圖4-11)。為了要把葡萄糖的6個碳原子一分為二,先使它兩端磷酸化,才能斷列成兩個接近相等的部分,然後脫去磷酸成丙酮酸。
上述反應都在細胞漿中進行。每一步都有特定的酶參加。第①、③兩步消耗2分子ATP,說明葡萄糖分解代謝開始時要用能量來推動。⑥、⑨兩步各產生2分子ATP。從葡萄到丙酮酸,淨生2分子ATP,酵解產生的能量不多。
細胞漿中NAD+和NADH的總量基本恆定。如所有的NAD+都變成NADH,酵解就得停止。要使葡萄繼續不斷氧化成酮酸,必需把NADH的氫轉移到另外的受體上去。有氧時氫經呼吸鏈氧化成水。無氧時氫交給丙酮酸使成乳酸⑩。乳酸過多時離開細胞入血。所以嚴重缺氧者可發生代謝性酸中毒。
(2)丙酮氧化成乙醯酶a 有氧時,丙酮酸進入粒線體。在丙酮酸脫氫酶系的幫助下,氧化脫羥成乙醯輔酸脫氫酶系的幫助下,氧化脫羥成乙醯輔酶A,同時產生1molATP和1molNADH。維生素B1B2、煙醯胺、泛酸參加。反應是不可逆的。
(3)三羧酸循環乙醯輔酶A上的兩個來自葡萄糖的碳原子,經三羥酸循環生成2molCO2,完成葡萄糖的氧化。反應在粒線體中進行。這一過程分步(圖4-12)。
三羧酸循環中產生3molNADH,1molFADH2,1molGTP。
NADH把氫通過FAD和細胞色素交給氧,同時放出能量。每molNADH通過這一系列反應所放出的能量是220kJ(52.6kCal)。這些能量以ATP的形式貯存起來。每molATP能產生30.5Kj(7.3kCal),理論上每molNADHRNUD氧化產生的能量可用來合成7molATP。這是指在標準狀況(25℃,1個大氣壓,作用物和產物濃度都是1M/LPH7)下說的。在模擬體內的條件時,實驗證明只能合成3molATP。如用反應式表示:
NADH+H++1/2O2
![]() NAD++H2O+能量
NAD++H2O+能量
3ADP+3H3PO4+能量
![]() 3ATP+3H2o
3ATP+3H2o
前面一個是氧化反應,後面一個是磷酸化,二者偶合在一起,稱為氧化磷酸化。
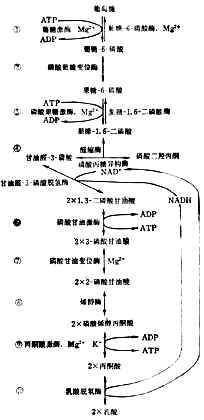
圖4-11 糖的酵解
實驗也證明FADH2氧化時產生2個ATP,GTP可生成1molATP。根據這些數值,葡萄糖完全氧化時,機體獲得的能量可歸納如下:
①酵解:葡萄糖→2丙酮酸+8ATp
②丙酮酸氧化成乙醯輔酶A:
2丙酮酸→2乙醯輔酶A+2CO2+6ATp
③三羧酸循環:
2乙醯輔酶A→4CO2+24ATp
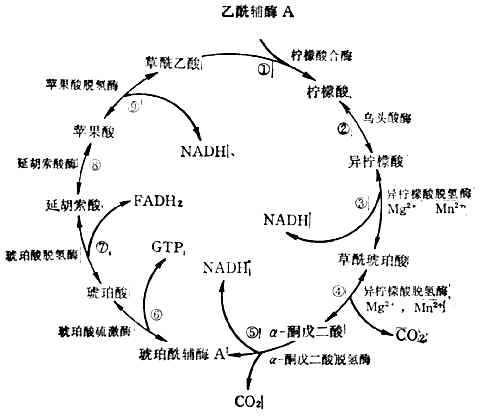
圖4-12三羧酸循環
即每mol葡萄糖在體內氧化成6molCO2時可產生38molATP,機體可以利用的能量是38×30或1140kJ[(38×7.3)或277kCal]。每mol葡萄糖氧化成6molCO2和水時,自由能的改變是2870KJ(686kCal)。所以,機械效率=1140/2870×10=40%。
這是利用標準狀況的數字計算的,在體內高一些,約60%。
從上面歸納的數字也可看出碳水化物提供的能量,在氧化的三個階段中,以三羧酸循環最多。脂肪和蛋白質最後也通過三羧酸循環氧化。食物向機體提供的能量,90%來自三羧酸循環。
葡萄糖代謝另外還有一種途徑,不通過果糖二磷酸,叫做已糖一磷酸通路。由於產生核榶,又叫做戊糖-磷酸通路。
(4)已糖-磷酸通路這一通路目的是提供合成核酸所需的核糖,以及合成脂肪酸、膽固醇等所需的能量供應者NADPH(還原型輔酶Ⅱ)(圖4-13)。肝臟中約有30%的葡萄糖經過這一通過代謝。紅細胞中這一通路也很活躍。它的失調可導致溶血性貧血。反應都在細胞漿中進行。
核糖-5-磷酸既可用於核酸的生物合成,又可與5-磷酸木酮糖反應生成甘油醛3-磷酸和果糖-6-磷酸。究竟走哪一條路,要看細胞本身對NADPH和核糖的需要量而定。許多細胞NADPH要得多,過剩的戊糖可經甘油醛-3-磷酸和果糖-6-磷酸酵解,或由果糖-6-磷酸轉變成葡萄糖-6-磷酸而重走這一通路。
這一通路需維生素B1參加。要使此路暢通,膳食中需有足量的維生素B1。5-磷酸核酮糖轉變為葡萄糖-6-磷酸的反應,可用於評價維生素B1參加。要使此路暢通,膳食中需有足量的維生素B1的營養狀況。
(5)糖原葡萄糖到各個細胞發揮作用以後,多餘的可儲存起來。避免細胞內滲透壓的升高,細胞把葡萄糖合成大分子的糖原。需要能量時糖原再分解葡萄糖。如肝糖原產生的葡萄糖,用以維持血糖的恆定。肌糖原產生的葡萄糖,直接用於供給能量。
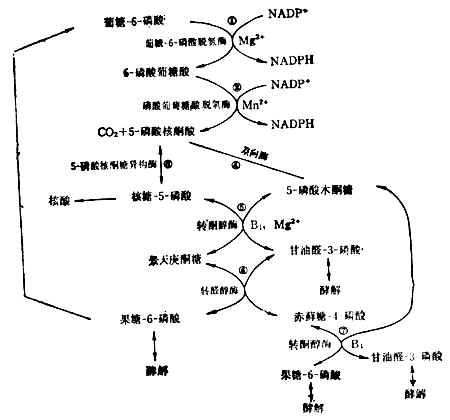
圖4-13 已糖-磷酸通路
4.2.3 三大營養代謝的關係
人習慣於三餐,飽腹和空腹交替產生。所以要具備能處理碳水貨物增多和缺少時保證能量供應的兩套本領。
餐後血糖升沿高,胰島素分泌增加,胰高血糖素分泌減少。使更多的葡萄糖進入肝臟、肌肉和脂肪組織;增強酶活力,加速葡萄的氧化和肝糖原、肌糖原的合成。超過糖原的儲存量後,肝臟可把葡萄糖經磷酸二羥丙酮還原成甘油-3-磷酸,與乙醯輔酶A合成的脂肪酸,利用NADPH提供的能量,生成脂肪。再和蛋白質結合成極低密度脂蛋白入血,運送到脂肪組織儲存。過多的碳水化物也能合成某些非必需胺基酸。
餐後約4h,不能利用食物中的葡萄糖。胰高血糖素分泌增加,胰島素減少,轉為利用肝糖原以供給血糖。肝糖原儲存量不多,成人能動用的不到70g,不夠一個晚上的消耗。所以,從餐後8h開始,還得利用脂肪以提供能量。甘油可變成糖,但在脂肪中所佔重量不多。脂肪酸只能供能,不能轉變為葡萄糖。因此,血糖的來源不得不轉向胺基酸了。一般說來,除亮氨酸和賴氨酸外,其他胺基酸都是能成糖的,但從能量的觀點看,只有丙氨酸和谷氨酸可作葡萄糖的來源。此時血糖的90%來自丙氨酸,10%來自谷氨酸。
肝臟能把肌肉中送來的丙氨酸作用成丙酮酸而生成葡萄糖。空腹時,肌肉中進入血液的胺基酸增加,其總量的30~40%是丙氨酸。但肌肉蛋白質的丙氨酸含量不到10%。另外的丙氨酸從哪裡來的?葡萄糖經酵解可生丙酮酸,再經轉氨作用丙氨酸。但這樣葡萄糖的量並沒有增加。增加的丙氨酸實際來自其他氨其酸。
腎臟把谷氨酸去氨後得α酮戊二酸,再和運來的成糖胺基酸如丙酸起轉氨作用而得丙酮酸,最後得葡萄糖。α-酮戊二酸也能經三羥酸循環形成蘋果酸,最後成葡萄糖。
| 關於「營養學/碳水化物的生化代謝」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |