生物化學與分子生物學/脂肪酸的合成
| 醫學電子書 >> 《生物化學與分子生物學》 >> 脂類代謝 >> 脂肪酸代謝 >> 脂肪酸的合成 |
| 生物化學與分子生物學 |
|
|
|
人體內的脂肪酸大部分來源於食物,為外源性脂肪酸,在體內可通過改造加工被人體利用。同時機體還可以利用糖和蛋白轉變為脂肪酸稱為內源性脂肪酸,用於甘油三酯的生成,貯存能量。合成脂肪酸的主要器官是肝臟和哺乳期乳腺,另外脂肪組織、腎臟、小腸均可以合成脂肪酸,合成脂肪酸的直接原料是乙醯CoA,消耗ATP和NADPH,首先生成十六碳的軟脂酸,經過加工生成人體各種脂肪酸,合成在細胞質中進行。
目錄 |
(一)軟脂酸的生成
脂肪酸的合成首先由乙醯CoA開始合成,產物是十六碳的飽和脂肪酸即軟酯酸(palmitoleic acid)。
1.乙醯CoA的轉移
乙醯CoA可由糖氧化分解或由脂肪酸、酮體和蛋白分解生成,生成乙醯CoA的反應均發生在粒線體中,而脂肪酸的合成部位是胞漿,因此乙醯CoA必須由粒線體轉運至胞漿。但是乙醯CoA不能自由通過粒線體膜,需要通過一個稱為檸檬酸丙酮酸循環(citratepyruvate cycle)來完成乙醯CoA由粒線體到胞漿的轉移。首先在粒線體內,乙醯CoA與草醯乙酸經檸檬酸合成酶催化,縮合生成檸檬酸,再由粒線體內膜上相應載體協助進入胞液,在胞液內存在的檸檬酸裂解酶(citrate lyase)可使檸檬酸裂解產生乙醯CoA及草醯乙酸。前者即可用於生成脂肪酸,後者可返回粒線體補充合成檸檬酸時的消耗。但草醯乙酸也不能自由通透粒線體內膜,故必須先經蘋果酸脫氫酶催化,還原成蘋果酸再經粒線體內膜上的載體轉運入粒線體,經氧化後補充草醯乙酸。也可在蘋果酸酶作用下,氧化脫羧生成丙酮酸,同時伴有NADPH的生成。丙酮酸可經內膜載體被轉運入粒線體內,此時丙酮酸可再羧化轉變為草醯乙酸。每經檸檬酸丙酮酸循環一次,可使一分子乙酸CoA由粒線體進入胞液,同時消耗兩分子ATP,還為機體提供了NADPH以補充合成反應的需要(見圖5-15)。
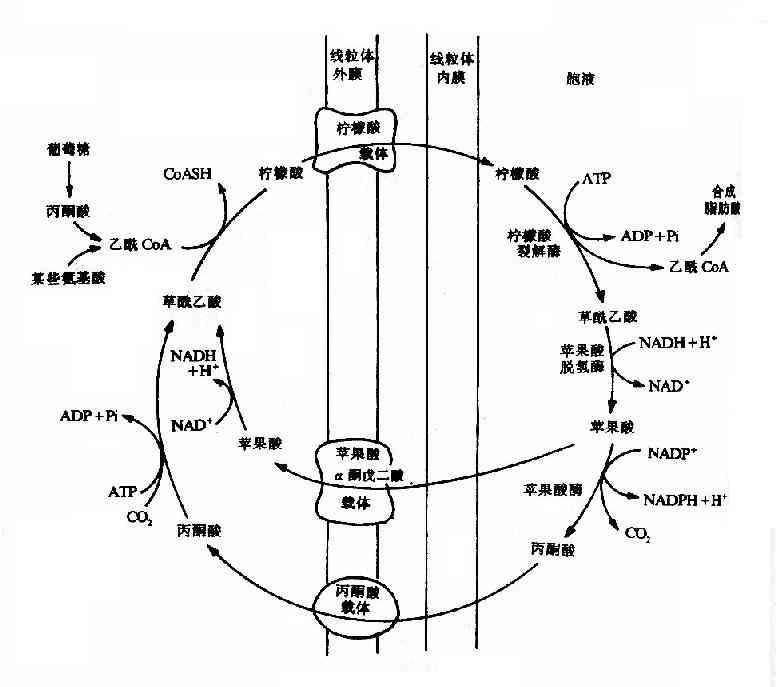
圖5-15 檸檬酸-丙酮酸循環
2.丙二醯CoA的生成
乙醯CoA由乙醯CoA羧化酶(acetyl CoA carboxylase)催化轉變成丙二醯CoA(或稱丙二酸單醯CoA)反應如下:

乙醯CoA羧化酶存在於胞液中,其輔基為生物素,在反應過程中起到攜帶和轉移羧基的作用。該反應機理類似於其他依賴生物素的羧化反應,如催化丙酮酸羧化成為草醯乙酸的反應等。
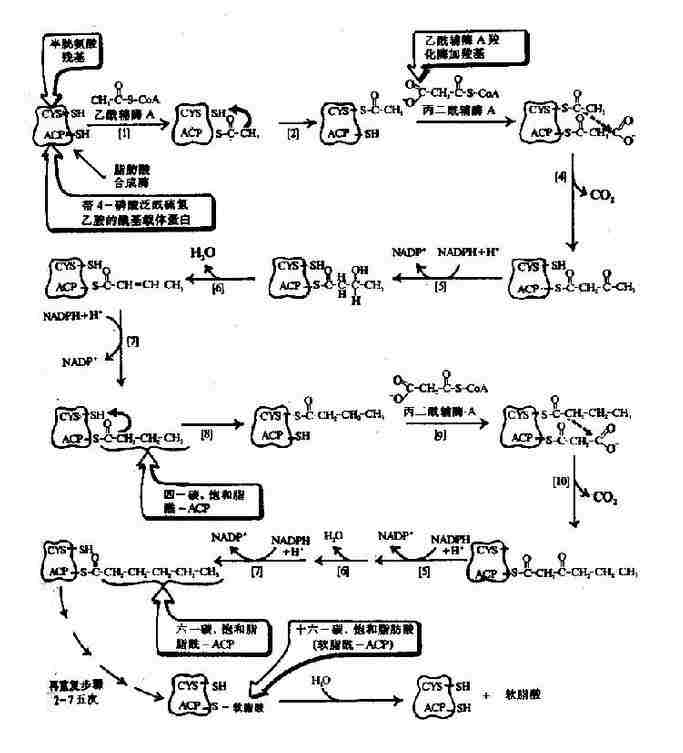
圖5-16 原核生物脂肪酸合成酶複合物生成軟脂酸(16:0)
由乙醯CoA羧化酶催化的反應為脂肪酸合成過程中的限速步驟。此酶為一別構酶,在變構效應劑的作用下,其無活性的單體與有活性的多聚體(由10?0個單體呈線狀排列)之間可以互變。檸檬酸與異檸檬酸可促進單體聚合成多聚體,增強酶活性,而長鏈脂肪酸可加速解聚,從而抑制該酶活性。乙醯CoA羧化酶還可通過依賴於cAMP的磷酸化及去磷酸化修飾來調節酶活性。此酶經磷酸化後活性喪失,如胰高血糖素及腎上腺素等能促進這種磷酸化作用,從而抑制脂肪酸合成;而胰島素則能促進酶的去磷酸化作用,故可增強乙醯CoA羧化酶活性,加速脂肪酸合成。
同時乙醯CoA羧化酶也是誘導酶,長期高糖低脂飲食能誘導此酶生成,促進脂肪酸合成;反之,高脂低糖飲食能抑制此酶合成,降低脂肪酸的生成。
3.軟脂酸的生成
軟脂酸的合成實際上是一個重複循環的過程,由1分子乙醯CoA與7分子丙二醯CoA經轉移、縮合、加氫、脫水和再加氫重複過程,每一次使碳鏈延長兩個碳,共7次重複,最終生成含十六碳的軟脂酸(圖5-16)。
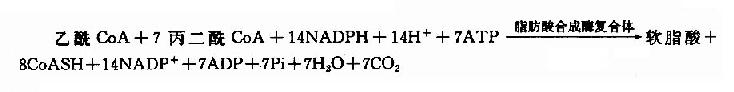
在原核生物(如大腸桿菌中)催化此反應的酶是一個由7種不同功能的酶與一種醯基載體蛋白(acyl carrier protein,ACP)聚合成的複合體。在真核生物催化此反應是一種含有雙亞基的酶,每個亞基有7個不同催化功能的結構區和一個相當於ACP的結構區,因此這是一種具有多種功能的酶。
脂肪酸合成需消耗ATP和NADPH+H+,NADPH主要來源於葡萄糖分解的磷酸戊糖途徑。此外,蘋果酸氧化脫羧也可產生少量NADPH。
脂肪酸合成過程不是β-氧化的逆過程,它們反應的組織,細胞定位,轉移載體,醯基載體,限速酶,激活劑,抑制劑,供氫體和受氫體以及反應底物與產物均不相同(表5-6)。
表5-6 脂肪酸合成和分解的比較
| 合成 | 分解 | |
| 反應最活躍時期 | 高糖膳食後 | 飢餓 |
| 刺激激素 | 胰島素/胰高血糖素高比值 | 胰島素/胰高血糖素低比值 |
| 主要組織定位 | 肝臟為主 | 肌肉、肝臟 |
| 亞細胞定位 | 胞漿 | 粒線體為主 |
| 醯基載體 | 檸檬酸(粒線體到胞漿) | 肉毒鹼(胞漿到粒線體) |
| 含磷酸醯疏基乙胺的活性載體 | 醯基載體蛋白區,CoA | CoA |
| 氧化還原輔因子 | NADPH | NAD+,FAD |
| 二碳供體/產物 | 丙二醯CoA;醯基供體 | 乙醯CoA:產物 |
| 激活劑 抑制劑 |
檸檬酸脂輔酶CoA(抑制乙醯CoA羧化酶) | 丙二醯CoA(抑制肉毒鹼醯基轉移酶) |
| 反應產物 | 軟脂酸 | 乙醯輔酶A |
(二)其它脂肪酸的生成
人體內不僅有軟脂酸,還有碳鏈長短不等的其它脂肪酸,也有各種不飽和脂肪酸,除營養必需脂肪酸依賴食物供應外,其它脂肪酸均可由軟脂酸在細胞內加工改造而成。
1.碳鏈的延長和縮短
脂肪酸碳鏈的縮短在粒線體中經β-氧化完成,經過一次β-氧化循環就可以減少兩個碳原子。
脂肪酸碳鏈的延長可在滑面內質網和粒線體中經脂肪酸延長酶體系催化完成。
在內質網,軟脂酸延長是以丙二醯CoA為二碳單位的供體,由NADPH+H+供氫,亦經縮合脫羧、還原等過程延長碳鏈,與胞液中脂肪酸合成過程基本相同。但催化反應的酶體系不同,其脂肪醯基不是以ACP為載體,而是與輔酶A相連參加反應。除腦組織外一般以合成硬脂酸(18C)為主,腦組織因含其他酶,故可延長至24碳的脂肪酸,供腦中脂類代謝需要。
在粒線體,軟脂酸經粒線體脂肪酸延長酶體系作用,與乙醯CoA縮合逐步延長碳鏈,其過程與脂肪酸β氧化逆行反應相似,僅烯脂醯CoA還原酶的輔酶為NADPH+H+與β氧化過程不同。通過此種方式一般可延長脂肪酸碳鏈至24或26碳,但以硬脂酸最多。
2.脂肪酸脫飽和
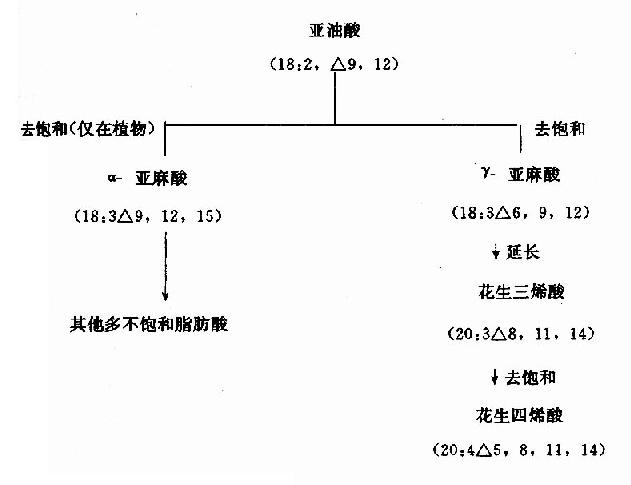
人和動物組織含有的不飽和脂肪酸主要為軟油酸(16:1△9)、油酸(18:1△9)、亞油酸(18:2△9,12)、亞麻酸(18:3△9,12,15)、花生四烯酸(20:4△5,8,11,14)等。其中最普通的單不飽和脂肪酸棗軟油酸和油酸可由相應的脂肪酸活化後經去飽和酶(acylCoAdesaturase)催化脫氫生成。這類酶存在於滑面內質網,屬混合功能氧化酶(見肝臟代謝章)。因該酶只催化在△9形成雙鍵,而不能在C10與末端甲基之間形成雙鍵,故亞油酸(linoleate)、亞麻酸(linolenate)及花生四烯酸(arachidonate)在體內不能合成或合成不足。但它們又是機體不可缺少的,所以必須由食物供給,因此,稱之為必需脂肪酸(essential fatty acid)。植物組織含有可以在C10與末端甲基間形成雙鍵(即ω3和ω6)的去飽和酶,能合成以上3種多不飽和脂肪酸。當食入亞油酸後,在動物體內經碳鏈加長及去飽和後,可生成花生四烯酸。
(三)脂肪酸合成的調節
乙醯CoA羧化酶催化的反應是脂肪酸合成的限速步驟,很多因素都可影響此酶活性,從而使脂肪酸合成速度改變。脂肪酸合成過程中其他酶,如脂肪酸合成酶、檸檬酸裂解酶等亦可被調節。
1.代謝物的調節
在高脂膳食後,或因飢餓導致脂肪動員加強時,細胞內軟脂醯CoA增多,可反饋抑制乙醯CoA羧化酶,從而抑制體內脂肪酸合成。而進食糖類,糖代謝加強時,由糖氧化及磷酸戊糖循環提供的乙醯CoA及NADPH增多,這些合成脂肪酸的原料的增多有利於脂肪酸的合成。此外,糖氧化加強的結果,使細胞內ATP增多,進而抑制異檸檬酸脫氫酶,造成異檸檬酸及檸檬酸堆積,在粒線體內膜的相應載體協助下,由粒線體轉入胞液,可以別構激活乙醯CoA羧化酶。同時本身也可裂解釋放乙醯CoA,增加脂肪酸合成的原料,使脂肪酸合成增加。
2.激素的調節
胰島素、胰高血糖素、腎上腺素及生長素等均參與對脂肪酸合成的調節。
胰島素能誘導乙醯CoA羧化酶、脂肪酸合成酶及檸檬酸裂解酶的合成,從而促進脂肪酸的合成。此外,還可通過促進乙醯CoA羧化酶的去磷酸化而使酶活性增強,也使脂肪酸合成加速。
胰高血糖素等可通過增加cAMP,致使乙醯CoA羧化酶磷酸化而降低活性,因此抑制脂肪酸的合成。此外,胰高血糖素也抑制甘油三酯合成,從而增加長鏈脂醯CoA對乙醯CoA羧化酶的反饋抑制,亦使脂肪酸合成被抑制。
(四)前列腺素、血栓素及白三烯
前列腺素(prostaglandin,PG),血栓素(thromboxane,TX)和白三烯(leukotrienes,LT)均由花生四烯酸衍生而來。它們在細胞內生成後,可作為調節物對幾乎所有的細胞代謝發揮調節作用,而且與炎症、過敏反應和心血管疾病等病理過程有關。
生物膜上的膜磷脂含有花生四烯酸,它可被磷脂酶A2水解,釋放花生四烯酸。花生四烯酸可在前列腺素內過氧化物合成酶催化下,消耗O2和還原性谷胱甘肽,發生氧化和環化反應,生成前列腺素H2。前列腺素H2可進一步衍生成其它前列腺素及血栓素(見圖5?7)。可的松(cortisol)抑制磷酸酶A2活性,減少花生四烯酸的生成,從而抑制前列原素的合成,阿斯匹林(aspirin)和保泰松(phenylbutazone)抑制前列腺素內過氧化物合成酶活性使前列腺素和血栓素生成減少。
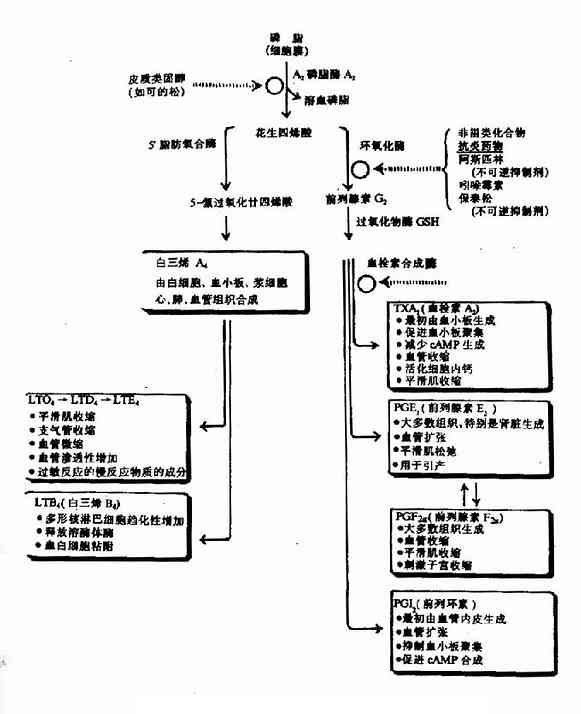
圖5-17 花生四烯酸生成PG,TX和LT概況
| 關於「生物化學與分子生物學/脂肪酸的合成」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |