醫學微生物學/細菌的結構
| 醫學電子書 >> 《醫學微生物學》 >> 細菌的形態與結構 >> 細菌的結構 |
| 醫學微生物學 |
|
|
細菌的結構對細菌的生存、致病性和免疫性等均有一定作用。細菌的結構按分布部位大致可分為:表層結構,包括細胞壁、細胞膜、莢膜;內部結構包括細胞漿、核蛋白體、核質、質體及芽胞等;外部附件,包括鞭毛和菌毛。習慣上又把一個細菌生存不可缺少的,或一般細菌通常具有的結構稱為基本結構,而把某些細菌在一定條件下所形成的特有結構稱為特殊結構(圖2-2)。
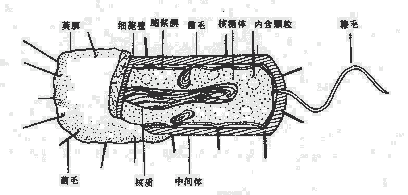
圖2-2 細菌的結構
一、基本結構
細菌基本結構包括細胞壁、細胞膜、細胞漿及核質。
(一)細胞壁(Cell wall)細胞壁為細菌表面比較複雜的結構。是一層較厚(5~80nm)、質量均勻的網狀結構,可承受細胞內強大的滲透壓而不破壞。細胞壁堅韌而有彈性。
1.細胞壁主要組份:主要成分是肽聚糖(Peptidoglycan),又稱粘肽(Mucopetide)。細胞壁的機械強度有賴於肽聚糖的存在。合成肽聚糖是原核生物特有的能力。肽聚糖是由N-乙醯葡萄糖胺和N-乙醯胞酸兩種氨基糖經β-1.4糖苷鍵連接間隔排列形成的多糖支架。在N-乙醯胞壁酸分子上連接四肽側鏈,肽鏈之間再由肽橋或肽鏈聯繫起來,組成一個機械性很強的網狀結構(圖2-3)。各種細菌細胞壁的肽聚糖支架均相同,在四肽側鏈的組成及其連接方式隨菌種而異。
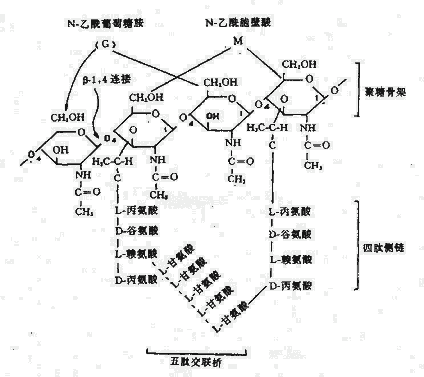
圖2-3 肽聚糖分子結構
革蘭氏陽性菌例如葡萄球的四肽側鏈胺基酸由D-丙-D-谷-r-L-賴-D-丙組成。初合成的肽鏈末端多一個D-丙氨酸殘基。肽橋是一條5個甘氨酸的肽鏈,交聯時一端與側鏈第三位上賴氨酸連接,另一端在轉肽酶的作用下,使另一條五肽側鏈末端D-丙氨酸脫去,而與側鏈第四位D-丙氨酸連接。從X光檢查可見肽聚糖的多糖鏈是一條較硬而又呈螺旋狀捲曲的長桿,由於其呈螺旋狀,連接在其上的肽鏈才伸向四方,使交聯受到一定了限制,只有鄰近的肽鏈才可交聯。但葡萄球菌的肽橋較長,有可塑性,使遠距離的肽鏈間也可交聯,交聯率達90%,形成堅固緻密的三維立本網狀結構(圖2-4)。
而革蘭氏陰性大腸桿菌的四肽側鏈中第三位的胺基酸被二氨基庚二酸(DAP)所取代,以肽鏈直接與相鄰四肽側鏈中的D-丙氨酸相連,且交聯率低,沒有五肽交聯橋,形成二維平面結構,所以其結構較革蘭氏陽性的葡萄球疏樺(圖2-5)。
|
|
|
| 圖2-4 金黃色葡萄球糖細胞壁的肽聚糖結構 | 圖2-5 大腸桿菌細胞壁的肽聚糖結構 |

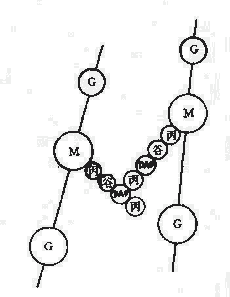
| M:N-乙醯胞酸 | G:N-乙醯氨基葡萄糖 | O:β-1,4糖苷鏈 | a:L-丙氨酸 |
| b:D-谷氨酸 | c:L-賴氨酸 | d:D-丙氨酸 | x:甘氨酸 |
凡能破壞肽聚糖結構或抑制其合成的物質,都能損傷細胞壁而使細菌變形或殺傷細菌,例如溶菌酶(Lysozyme)能切斷肽聚糖中N-乙醯葡萄糖胺和N-乙醯胞壁酸之間的β-1.4糖苷鍵之間的聯苷鍵之間的聯結,破壞肽聚糖支架,引起細菌裂解。青黴素和頭孢菌素能與細菌競爭合成胞壁過程所需的轉肽酶,抑制四肽側鏈上D-丙氨酸與五肽橋之間的聯結,使細菌不能合成完整的細胞壁,可導致細菌死亡。人和動物細胞無細胞壁結構,亦無肽聚糖,故溶菌酶和青黴素對人體細胞均無毒性作用。除肽聚糖這一基本成份以外,革蘭氏陽性菌和革蘭氏陰性菌還各有其特殊結構的成分。
2.革蘭氏陽性菌細胞壁特殊組份細胞壁較厚,約20~80mm。肽聚糖含量豐富,有15~50層,每層厚度1nm,約佔細胞壁乾重的50~80%。此外,尚有大量特殊組份磷壁酸(Teichoic acid)。 (圖2-6)磷壁酸是由核糖醇(Ribitol)或甘油(Glyocerol)殘基經由磷酸二鍵互相連接而成的多聚物。磷壁酸分壁磷壁酸(Wallteichoic acid)和膜磷壁酸(Membrane teichoic acid)兩種,前者和細胞壁中肽聚糖的N-乙醯胞壁酸連結,膜磷壁酸又稱脂磷壁酸(Lipteichoic acid)和細胞膜連結,另一端均游離於細胞壁外。磷壁酸抗原性很強,是革蘭氏陽性菌的重要表面抗原;在調節離子通過粘肽層中起作用;也可能與某些酶的活性有關;某些細菌的磷壁酸,能粘附在人類細胞表面,其作用類似菌毛,可能與致病性有關。
此外,某些革蘭氏陽性菌細胞壁表面還有一些特殊的表面蛋白,如A蛋白等,都與致病有關。
3.革蘭氏陰性菌細胞壁特殊組份細胞壁較薄,約10~15nm,有1~2層肽聚糖外,約佔細胞壁乾重的5~20%。結構比較複雜。尚有特殊組份外膜層位於細胞壁肽聚糖層的外側,包括脂多糖、脂質雙層、脂蛋白三部分(圖2-6)。
脂蛋白(Lipoprotein)一端以蛋白質部分共價鍵連接於肽聚糖的四肽側鏈上另一端以脂質部分經共價鍵連接於外膜的磷酸上。其功能是穩定外膜並將之固定於肽聚糖層。
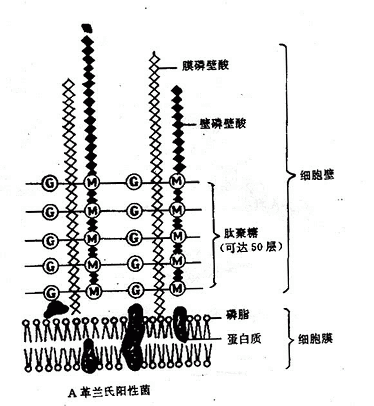
圖2-6 細菌細胞壁結構模式圖
脂質雙層是革蘭陰性菌細胞壁的主要結構,除了轉運營養物質外,還有屏障作用,能阻止多種物質透過,抵抗許多化學藥物的作用,所以革蘭氏陰性菌對溶菌酶、青黴素等比革蘭氏陽性具有較大的抵抗力。一些化學物質如乙二胺四乙酸(EDTA)與2%十二烷基硫酸鈉(SDS)或45%酚水溶液可以將外膜除去,而留下堅韌的肽聚糖層。此外,外膜蛋白質還可作為某些噬菌體和性菌毛的受體。
脂多糖(Lipopolysacchride,LPS)由脂質雙層向細胞外伸出,包括類脂A、核心多糖、特異性多糖三個組成部分,習慣上將脂多糖稱為細菌內毒素。
①類脂A:為一種糖磷脂,是由焦磷酸鍵聯結的氨基葡萄糖聚二糖鏈,其上結合有各種長鏈脂肪酸。它是脂多糖的毒性部分及主要成份。為革蘭氏陰性菌的致病物質。無種屬特異性,各種革蘭氏陰性菌內毒性引起的毒性作用都大致相同。
②核心多糖:位於類脂A的外層,由已糖、瘐糖、2-酮基—3-脫氧辛酸(KDO)、磷酸乙醇胺等組成。經KDO與類質A共價聯結。核心多糖具有屬特異性,同一屬細菌的核心多糖相同。
③特異性多糖:在脂多糖的最外層,是由數個至數十個低聚糖(3~5單糖)重複單位所構成的多糖鏈。革蘭氏陰性菌的菌體抗原(O抗原)就是特異多糖。各種不同的革蘭氏陰性菌的特異性多糖種類及排列順序各不相同,從而決定了細菌抗的特異性。
革蘭氏陽性菌和革蘭氏陰性菌的細胞壁結構顯著不同,導致這兩類細菌在染色性、抗原性、毒性、對某些藥物的敏感性等方面的很大差異。
表2-1 革蘭氏陽性菌與革蘭氏陰性菌細胞壁結構的比較
| 特 征 | 革蘭氏陽性菌 | 革蘭氏陰性菌 |
| 強度 | 較堅韌 | 較疏鬆 |
| 厚度 | 厚,20~80nm | 薄,5~10nm |
| 肽聚糖層數 | 多,可達50層 | 少,1~3層 |
| 肽聚糖含量 | 多,可占胞壁乾重50~80% | 少,占胞壁乾重10~20% |
| 磷壁酸 | + | - |
| 外膜 | - | + |
| 結構 | 三維空間(立體結構) | 二維空間(平面結構) |
4.細胞壁的功能細菌細胞壁堅韌而富有彈性,保護細菌抵抗低滲環境,承受世界盃內的5~25個大氣的滲透壓,並使細菌在低滲的環境下細胞不易破裂;細胞壁對維持細菌的固有形態起重要作用;可充許水分及直徑小於1nm的可溶性小分子自由通過,與物質交換有關;細胞壁上帶有多種抗原決定簇,決定了細菌菌體的抗原性。
5.L型細菌 L型是指細菌發生細胞壁缺陷的變型。因其首次在Lister研究所發現。故以其第一個字母命名。當細菌細胞壁中的肽聚糖結構受到理化或生物因素的直接破壞或合成被抑制這種細胞壁受損的細菌一般在普通環境中不能耐受菌體內部的高滲透壓而將脹裂死亡;但在高滲環境下,它們仍可存活而成為細菌細胞壁缺陷型。革蘭氏陽性菌L型稱為原生質體(protoplast),必須生存於高滲環境中。革蘭氏陰性菌L型稱為原生質球(spheroplast),在低滲環境中仍有一定的抵抗力。
細菌L型的形態因缺失細胞壁而呈高度多形性,有球狀、桿狀和絲狀。大小不一,L型細菌大多數染成革蘭氏陰性。細菌L型生長繁殖時的營養要求基本與原菌相同,但必須補充3%~5%的NaC1、10%~20%蔗糖或7%聚乙烯吡咯酮(PVP)等穩定劑,以提高培養基的滲透壓。同時還需要加入人或馬血清。L型細菌生長較緩慢,一般培養2~7天後在軟瓊脂平板形成中同較厚、四周較薄的荷包蛋樣細小菌落。此外,L型菌尚有顆粒型和絲狀型兩種類型。L型細菌在液體培養基中生長後呈較疏鬆的絮狀顆粒,沉於管底,培養液則澄清。
人工誘導或自然情況下,細菌L型在體內或體外均能產生。各種細菌L型有一個共同的致病特點。即引起多組織的間質性炎症。細菌變為L型致病性有所減弱,但在一定條件下L型又可復為細菌型,引起病情加重。變形後的細菌其形態、培養特性均發生了改變,以致查不出病原使許多病人貽誤診治。臨床遇有症状明顯而標本常規細菌培養陰性者,應考慮細菌L型感染的可能性,宜作細菌L型的專門培養。
(二)細胞膜(Cellmembrane)或稱胞膜(Cytoplasmic membrane)位於細胞壁內側,包繞在細菌胞漿外的具有彈性的半滲透性脂質雙層生物膜。主要由磷脂及蛋白質構成,膜不含膽固醇是與真核細胞膜的區別點。細胞膜有選擇性通透作用,與細胞壁共同完成菌體內外的物質交換。膜上有多種呼吸酶,參與細胞的呼吸過程。膜上有多種合成酶,參與生物合成過程。細菌細胞膜可以形成特有的結構。
1.中介體(Mesosome)用電子顯微鏡觀察,可以看到細胞膜向胞漿凹陷摺疊成囊狀物,稱為中介體。中介體與細胞的分裂、呼吸、胞壁合成和芽胞形成有關。中介體位置常在菌體的側面或靠近中央橫隔處。橫隔中介體與核質相連,當細菌分裂時橫隔中介體也一分為二,各自帶一套核質進入子代細胞;中介體擴大了細胞膜的表面積,相應地增加呼吸酶的含量,可為細菌提供大量能量,有擬粒線體(Chondroid)之稱,中介體多見於革蘭氏陽性菌。
2.胞質間間隙在革蘭氏陰性細菌的細胞膜與細胞壁之間有一空間,稱為胞質間間隙(Periplasmic space)。此處聚集了若干種胞外酶,主要是水解酶,與營養物質的分解、吸收和運轉有關。能破壞某些抗生素的酶(如青黴素酶)亦集中在此間隙內。
(三)胞漿(Cytoplasm)是無色透明膠狀物,基本成份是水、蛋白質、脂類、核酸及少量無機鹽。細胞漿中還存在一些胞漿顆粒。
1.質體(Plasmid)這是染色體外的遺傳物質,為雙股環狀DNA。分子量比染色體小,可攜帶某些遺傳信息,例如耐藥因子、細菌素及性菌毛的基本均編碼在質體上。質體能進行獨立複製,失去質體的細菌仍能正常存活。質體可通過接合、轉導作用等將有關性狀傳遞給另一細菌。
2.核糖體(Ribosome)電鏡下可見到胞漿中有大量沉降係數為70S的顆粒,即核糖體。其化學組成70%為RNA,30%為蛋白質。細胞中約90%的RNA和40%的蛋白質存在於核糖體中。當mRNA連成多聚核蛋白體(Polyribosome),就成為合成蛋白質的場所。細菌的70S核糖體由50S和30S兩個亞基組成。鏈黴素能與細菌核糖體的30S基結合,紅霉素能與50S亞基結合,從而干擾細菌蛋白質的合成而導致細菌的死亡;真核細胞的核糖體為80S,因此對人體細胞則無影響。
3.胞漿顆粒(Cytoplasmagranula)大多數為營養貯藏物,較為常見的是貯藏高能磷酸鹽的異染顆粒(Metachrometicgranula),嗜鹼性較強,用特殊染色法可以看得更清晰。根據異染顆粒的形態及位置,可以鑒別細菌。
(四)核質(Nnclearmateral)或擬核(Nucleoid)是細菌的遺傳物質,決定細菌的遺傳特徵。集中在細胞漿的某一區域,多在菌體中部。它與真核細胞的細胞核不同點在於四周無核膜,故不成形,也無組蛋白包繞。一個菌體內一般含有1~2個核質。現已證明,細菌的核質是由雙股DNA組成的單一的一根環狀染色體反覆迴旋盤繞而成,細菌的染色體是裸露的DNA。
大腸桿菌的染色體分子量為3×109,伸展後長度約達1.1mm,約含5×106鹼基對,足可攜帶3,000~5,000個基因,以滿足細菌生命活動的全部需要,核質具有細胞核的功能,控制細菌的各種遺傳性狀。細菌胞漿中含有大量RNA,用鹼性染料染色著色很深,將核質掩蓋,不易顯露。若用酸或RNA酶處理,使RNA水解,再用富爾根(Feulgen)氏法染色,便可染出核質,在普通光學顯微鏡下可以看見,一般呈球狀、棒狀或啞鈴狀。
二、特殊結構
細菌的特殊結構包括莢膜、鞭毛、菌毛和芽胞。
(一)莢膜(Capsule)許多細菌胞壁外圍繞一層較厚的粘性、膠凍樣物質,其厚度在0.2um以上,普通顯微鏡可見,與四周有明顯界限,稱為莢膜。如肺炎雙球菌(圖2-7)。其厚度在0.2um以下者,在光學顯微鏡下才不能直接看到,必須以電鏡或免疫學方法才能證明,稱為微莢膜(Microcapsule),如溶血性鏈球菌的M蛋白、傷寒桿菌的Vi抗原及大腸桿菌的K抗原等。

圖2-7 肺炎球菌莢膜
大多數細菌(如肺炎球菌、腦膜炎球菌等)的莢膜由多糖組成。鏈球菌莢膜為透明質酸;少數細菌的莢膜為多肽(如炭疽桿菌莢膜為D-谷氨酸的多肽)。
細菌一般在機體內和營養豐富的培養基中才能形成莢膜。有莢膜的細菌在固體培養基上形成光滑型(S型)或粘液型(M)菌落,失去莢膜後菌落變為粗糙型(R)。莢膜並非細菌生存所必需,如莢膜丟失,細菌仍可存活。
莢膜除對鑒別細菌有幫助外,還能保護細菌免遭吞噬細胞的吞噬和消化作用,因而與細菌的毒力有關。莢膜抗吞噬的機理還不十分清楚,可能由於莢膜粘液層比較光滑,不易被吞噬細胞捕捉之故。莢膜能貯留水分使細菌能抗乾燥,並對其他因子(如溶菌酶、補體、抗體、抗菌藥物等)的侵害有一定抵抗力。
(二)鞭毛(Flagllum)在某些細菌菌體上具有細長而彎曲的絲狀物,稱為鞭毛。鞭毛的長度常超過菌體若干倍。不同細菌的鞭毛數目、位置和排列不同,可分為單毛菌(Monotrichate)、雙毛菌(Amphitrichate)、絲毛菌(Lophotrichate)、周毛菌(Peritrichate)(圖2-8)。

圖2-8 細菌的鞭毛(示意)
鞭毛自細胞膜長出,游離於細胞外。用電子顯微鏡研究鞭毛的超微結構,發現鞭毛的結構分為:基礎小體、鉤狀體和絲狀體三個部分組成(圖2-9)。
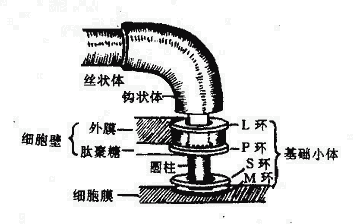
圖2-9 大腸桿菌鞭毛根部結構模式圖
1.基礎小體(Basalbody)位於鞭毛根部,埋在細胞壁中。革蘭氏陰性菌鞭毛的基礎小體由一根圓柱和兩對同心環所組成,一對是M環與S環,附著在細胞膜上;另一對是P環與L環,連在胞壁的肽聚糖和外膜上(M、S、P、L分別代表細胞膜、膜上、肽聚糖、外膜中的脂多糖)。革蘭氏陽性菌的細胞壁無外膜,其鞭毛只有M與S環而無P環和L環。鞭毛運動需要能量,細胞膜中的呼吸鏈可供其所需。
2.鉤狀體(Hook)位於鞭毛伸出菌體之處,呈鉤狀彎曲,鞭毛此轉變向外伸出,成為絲狀體。
3.絲狀體(Filament)呈纖絲狀,伸出於菌體之外,是由鞭毛蛋白亞單位呈緊螺旋狀纏繞而成的中空的管狀結構。鞭毛蛋白是一種纖維蛋白,其胺基酸組成與骨骼肌動蛋白相似,可能與鞭毛的運動性有關。
鞭毛是細菌的運動器官,往往有化學趨向性,常朝向有高濃度營養物質的方向移動,而避開對其在害的環境。常存在於桿菌及弧菌中。鞭毛的數量、分布可用以鑒別細菌。鞭毛抗原有很強的抗原性,通常稱為H抗原,對某些細菌的鑒定、分型及分類具有重要意義。
(三)菌毛(Pilus)菌毛是許多革蘭氏陰性菌菌體表面遍布的比鞭毛更為細、短、直、硬、多的絲狀蛋白附屬器,也叫做纖毛(Fimbriae)。其化學組成是菌毛蛋白(Pilin),菌毛與運動無關,在光鏡下看不見,使用電鏡才能觀察到。菌毛可分為普通菌毛(Commonpilus)和性菌毛(Sexpilus)兩種。
1.普通菌毛 長0.3~1.0um,直徑7nm。具有粘著細胞(紅細胞、上皮細胞)和定居各種細胞表面的能力,它與某些細菌的的致病性有關。無菌毛的細菌則易被粘膜細胞的纖毛運動、腸蠕動或尿液沖洗而被排除,失去菌毛,致病力亦隨之喪失。
2.性菌毛 有的細菌還有1~4根較長的性菌毛,比普通菌毛而粗,中空呈管狀。性菌毛由質體攜帶的一種致育因子(Ferilityfactor)的基因編碼,故性菌毛又稱F菌毛。帶有性菌毛的細菌稱為F+菌或雄性菌,無菌毛的細菌稱為F-菌或雌性菌。性菌毛能在細菌之間傳遞DNA,細菌的毒性及耐藥性即可通過這種方式傳遞,這是某些腸道桿菌容易產生耐藥性的原因之一。
(四)芽胞(Spore)在一定條件下,芽胞桿菌屬(如炭疽桿菌)及梭狀芽胞桿菌屬(如破傷風桿菌、氣性壞疽病原菌)能在菌體內形成一個折光性很強的不易著色小體,稱為內芽胞(Endospore),簡稱芽胞。
芽胞一般只在動物體外才能形成,並受環境影響,當營養缺乏,特別是碳源、氮源或磷酸鹽缺乏時,容易形成芽胞。不同細菌開成芽胞還需不同的條件,如炭疽桿菌須在有氧條件下才能形成芽胞。成熟的芽胞可被許多正常代謝物如丙氨酸、腺苷、葡萄糖、乳酸等激活而發芽,先是芽胞酶活化,皮質層及外殼迅速解聚,水分進入,在合適的營養和溫度條件下,芽胞的核心向外生長成繁殖體,開始發育和分裂繁殖。芽胞並非細菌的繁殖體,而是處於代謝相對靜止的休眠休態,以維持細菌生存的持久體。
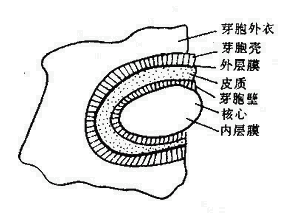
芽胞含水量少(約40%),蛋白質受熱不易變性。芽胞具有多層厚而緻密的胞膜,由內向外依次為核心、內膜、芽胞壁、皮質、外膜、芽胞殼和芽胞外衣(圖2-10)。特別是芽胞殼,無通透性,有保護作用,能阻止化學品滲入。芽胞形成時能合成一些特殊的酶,這些酶較之繁殖體中的酶具有更強的耐熱性。芽胞核心和皮質層中含有大量吡啶二羧酸(Dipicolinic acid,DPA),占芽胞乾重的5~15%,是芽胞所特有的成分,在細菌繁殖體和其他生物細胞中都沒有。DPA能以一種現尚不明的方式,使芽胞的酶類具有很高的穩定性。芽胞形成過程中很快合成DPA,同時也獲得耐熱性。
|
|
|
| 圖2-10 芽胞結構模式 | 圖2-11 細菌的芽胞形態 |
芽胞呈圓形或橢圓形,其直徑和在菌體內的位置隨菌種而不同,例如,炭疽桿菌的芽胞為卵圓形、比菌體小,位於菌體中央;破傷風桿菌芽胞正圓形、比菌體大,位於頂端,如鼓槌狀。這種形態特點有助於細菌鑒別(圖2-11)。芽胞在自然界分布廣泛,因此要嚴防芽胞污染傷口、用具、敷料、手術器械等。芽胞的抵抗力強,對熱力、乾燥、輻射、化學消毒劑等理化因素均有強大的抵抗力,用一般的方法不易將其殺死。有的芽胞可耐100℃沸水煮沸數小時。殺滅芽胞最可靠的方法是高壓蒸汽滅菌。當進行消毒滅菌時往往以芽胞是否被殺死作為判斷滅菌效的指標。
| 關於「醫學微生物學/細菌的結構」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |