生理學/脊髓對軀體運動的調節
| 醫學電子書 >> 《生理學》 >> 神經系統 >> 神經系統對軀體運動的調節 >> 脊髓對軀體運動的調節 |
| 生理學 |
|
|
|
在脊髓的前角中,存在大量運動神經元(α和γ運動神經元),它們的軸突經前根離開脊髓後直達所支配的肌肉。α運動神經元的大小不等,胞體直徑從幾十到150μm;大α運動神經元支配快肌纖維,小α運動神經地支配慢纖維。α運動神經元接受來自皮膚、肌肉和關節等外周傳入的信息,也接受從腦幹到大腦皮層等主位中樞傳的信息,產生一定的反射傳出衝動。因此,α運動神經元是軀幹骨骼肌運動反射的最後公路。
α運動神經元的軸突在離開脊髓走和肌肉時,其末梢在肌肉中分成許多小支,每一小支支配一根骨骼肌纖維。因此,在正常情況下,當這一神經元發生興奮時,興奮可傳導到受它支配的許多肌纖維,引起其收縮。由一個α運動神經元及其支配的全部肌纖維所組成的功能單位,稱為運動單位。運動單位的大小,決定於神經元軸突開梢分支數目的多少,一般是肌肉愈大,運動單位也愈大。例如,一個眼外肌運動神經元只支配6-12根肌纖維,而一個肢肌(如三角肌)的運動神經元所支配的肌纖維數目可達2000根。前者有利於肌肉進行精細的運動,後者有利於產生巨大的肌張力。同一個運動單位的肌纖維,可以和其他運動單位的肌纖維交叉分布,使其所佔有的空間範圍比該單位肌纖維截而的總和大10-30倍;因此,即使只有少數運動神經元活動,在肌肉中產生的張力也是均勻的。
γ運動神經元的胞體分散在α運動神經元之間,其胞體較α運動神經元為小。γ運動神經元的軸突也經前根離開脊髓,支配骨骼肌骨的梭內肌纖維。據觀察,前根中神經纖維的三分來自γ運動神經元。γ運動神經元的興奮性較高,常以較主頻率持續放電。在安靜和麻醉的運動中都觀察到,即使α運動神經元無放電,一些γ運動神經元仍持續放電。γ運動神經元和α運動神經元一樣,末梢也是釋放乙醯膽鹼作為遞質的。在一般情況下,當α運動神經元活動增加時,γ運動神經元也相應增加,從而調節著肌梭對牽拉刺激的敏感性。
目錄 |
(一)脊休克
在動物中將脊髓與延髓的聯繫切斷,用以研究脊髓單獨的功能;但為了保持動物的呼吸功能,常在頸脊髓第五節水平以下切斷,以保留膈神經對膈肌呼吸的傳出支配。這種脊髓與高位中樞離斷的動物稱為脊動物。在脊動物可以觀察到脊髓的一些基本功能;但由於失去了高位中樞的調節,因而不能完全反映正常的脊髓功能。
與高位中樞離斷的脊髓,在手術後暫時喪失反射活動的能力,進入無反應狀態,這種現象稱為脊休克(spinal shock)。脊休克的主要表現為:在橫斷面以下的脊髓所支配的骨骼肌緊張性減低甚至消失,血壓下降,外周血管擴張,發汁反射不出現,坡腸和膀胱中糞尿積聚,說明動物軀體與內臟反射活動均減退以至消失。脊休克現象只發生在切斷水平以下的部分。以後,一些以脊髓為中樞的反射活動可以逐漸恢復,恢復的迅速與否,與動物種類有密切關係;低等動物如蛙在脊髓離斷後數分鐘內反射即恢復,在犬則需幾天,而在人類則需數周以至數月(人類由於外傷等原因也可出現脊休克)。顯然,反射恢復的速度與不同動物脊髓反射依賴於高位中樞的程度有關。反射恢復過程中,首先是一引起比較簡單、比較原始的反射先恢復,如屈肌反射、腱反射等;然後才是比較複雜的反射逐漸恢復,如對側伸肌反射、搔爬反射等。反射恢復後的動物,血壓也逐漸上升到一定水平,動物可具有一定的排糞與排快活反射,說明內臟反射活動也能部分地恢復。反射恢復後,有些反射反應比正常時加強並廣泛擴散,例如屈肌反射、發汗反射等。
脊休克的產生並不由於切斷損傷的刺激性影響引起的,因為反射恢復後進行第二次脊髓切斷損務並不能使脊休克重現。所以,疹休克的產生原因是由於離斷的脊髓突然失去了高位中樞的調節,這裡主要指大腦皮層、前庭核和腦幹網狀結構的下行纖維對脊髓的易化作用。
脊休克的產生與恢復,說明脊髓可以完成某些簡單的反射活動,但正常時它們是在高位中樞調節下進行活動的。高位中樞對脊髓反射既有易化作用的一面,也有抑制作用的一面。例如,切斷脊髓後伸肌反射往往減弱,說明高位中樞對脊髓伸肌反射中樞有易化作用;而發汗反射加強,又說明高位中樞對脊髓發汗反射中樞有抑制作用。
脊動物由於脊髓離斷,脊髓內上行與下行的神經束秀難重新接通,造成感覺傳入衝動不能上達大腦皮層,而大腦皮層的傳出衝動也不能下達脊髓,以致在離斷水平以上就失去了知覺和所謂隨意動作。中樞的神經軸突與外周的神經軸突都有再生能力,但由於脊髓離斷部位有大量膠質細胞浸潤並逐步形成瘢痕,使軸突再生受阻。
脊髓離斷後屈肌反射比正常時加強,而伸肌反射往往減弱,以致屈肌反射常佔優勢,這不利於癱瘓肢體支持體重。因此,在低位脊髓橫貫性損傷的病人,通過站立姿勢的積極鍛煉以發展伸肌反射是很重要的;這種鍛煉使下肢伸肌具有足夠的緊張性以保持伸直,以使不依靠拐杖站立或行走。同時,通過鍛煉充分發揮末癱瘓肌肉的功能,例如背闊肌等由脊髓離斷水平以上的神經所支配,而卻隨著於骨盆,這樣就有可能使病人在借拐杖行走時擺動骨盆。
(二)屈肌反射與對側伸肌反射
在脊動物的皮膚接受傷害性刺激時,受刺激一側的肢體出現屈曲的反應,關節的屈肌收縮而伸肌馳緩,稱為屈肌反射。屈肌反射具有保持性意義。屈肌反射的強度也刺激強度有關,例如足部的較弱刺激只引致踝關節屈曲,刺激強度加大,則膝關節及髖關節也可發生屈曲。如刺激強度更大,則可以同側肢體發生屈肌反向的基礎上出現對側肢體伸直的反射活動,稱為對側伸肌反射。對側伸肌反射是姿勢反射的之一,具有維持姿勢的生理意義,動物一側肢體屈曲,對側肢體伸直以支持體重。屈肌反射是一種多突觸反射,其反射弧傳出部分可通向許多關節的肌肉。
在人類由於錐體束或大腦皮層運動區的功能發生障礙,脊髓失去了運動區的調節,可出現一處特殊的反射。例如,以鈍物劃足跖外側時,出現大趾背屈,其他四趾向外展開如扇形的反射,稱為巴彬斯基征(Babinski』s sign)陽性。從生理學角度來看,這一反射屬於屈肌反射,因為當刺激加強時還可伴有踝、膝、髖關節的屈曲。平時脊髓在大腦皮層的調節下,這一原始的屈肌反射被抑制而不表現出來。在嬰兒的錐體束未發育完全以前,以及成人深睡或麻醉狀態下,也可以出現巴彬斯基征陽性。
(三)牽張反射
有神經支配的骨骼肌,如受到外力牽拉使其伸長時,能產生反射效應,引起受牽扯的同一肌肉收縮,此稱為牽張反射(stretch reflex)。牽張反射有兩種類型,一種為腱反射,也稱位相性牽張反射;另一種為肌緊張,也稱緊張性牽張反射。
腱反射是指快速牽拉肌腱時發生的牽張反射。例如,叩擊膝關節下的股四頭肌腱使之受到牽扯,則股四頭肌即發生一次收縮,這稱為膝反射;叩擊跟腱使之受到牽扯,則小腿腓腸肌即發生一次收縮,這稱為跟腱反射。這些腱反射的感受器為肌梭,傳入神經纖維的直徑較粗(12-20μm)、傳導速度較快(90m/s以上),效應器為同一肌肉的肌纖維;反射反應的潛伏期很短,據測算興奮通過中樞的傳布時間公0.7ms左右,只夠一次突觸接替的中樞延擱時間。因此,腱反射為單突觸反射,傳入神經纖維經背根進入脊髓灰質後,直達前角與運動神經元發生突觸聯繫。當叩擊肌腱時,肌肉內的肌梭同時受到牽張,同時發動牽張反射。因此肌肉的收縮幾乎是一次同步性收縮。腱反射主要發生於肌肉內收縮較快的快肌纖維成分。
肌緊張是指緩慢持續牽拉腱時發生的牽張反射,其表現為受牽拉攏肌肉能發生緊張性收縮,阻止被拉長。肌緊張是維持軀體姿勢最基本的反射活動,是姿勢反向的基礎。例如,由於重力影響,支持體重的關節趨向於被重力所彎曲,關節彎曲必使伸肌肌腱受到持續牽拉,從而產生牽張反射引起該肌的收縮,對抗關節的屈曲,維持站立姿勢。由於重力經常作用於關節,因此這種牽張反射也就持續著。肌緊張與腱反射的反射弧基本相似,感受器也是肌梭,但中樞的突觸接替可能不止一個,即可能是多突觸反射,其效應器主要是肌肉內收縮較慢的慢肌纖維成分。肌緊張的反射收縮力量並不大,只是抵抗肌肉被牽拉,因此不表現明顯的動作。這可能是因為在同肌肉內的不同運動單位進行交替性的收縮而不是同步性收縮,所以肌緊張能持久維持而不易疲勞。
牽張反射主要是使受牽拉的肌肉發生收縮,但同一關節的協同肌也能發生興奮,而同一關節的頡頏肌則受到抑制(交互抑制),但並不影響其它關節的肌肉,伸肌和屈肌都有牽張反射,但脊髓的牽張反射主要表現在伸肌。屈肌的牽張反射表現不明顯,主要表現為它的頡頏肌(即伸肌)受到抑制。牽張反射(尤其是肌緊張)的主要生理意義在於維持這種站立姿勢,因此伸肌比屈肌的牽張反射明顯符合生理情況。
牽張反射的基本反射弧是簡單的,但在整體內牽張反射是受高位中樞調節的,而且可以建立條件反射。腱反射的減弱或消失,常提示反射弧的傳入、傳出通路受脊髓反射中樞的損害或中斷;而腱反射的亢進,則常提示高位中樞的病變I(例如錐體束症候群)。因此,臨床上常用測定腱反射的方法來了解神經系統的功能狀態。常用的腱反射見表10-5。
表10-5 常用的腱反射
| 名稱 | 檢查方法 | 中樞部位 | 效應 | |
| 肘反射 | 扣擊肱二頭肌肌腱 | 頸5-7 | 肘部屈曲 | |
| 膝反射 | 扣擊髕韌帶 | 腰2-4 | 小腿伸直 | |
| 跟腱反射 | 扣擊跟腱 | 腰5-骶2 | 腳向足底方向屈曲 | |
肌梭和腱器官
肌梭是一種感受肌肉長度變化或感受牽拉刺激的特殊的梭形感受裝置,長几個毫米,外層為一結諦組織囊(圖10-30)。肌梭囊內一般含有6-12根肌纖維,稱為梭內肌纖維;而囊外的一般肌纖維就稱為梭外肌纖維。整個肌梭現階附著在梭外肌纖維上,並與其平等排列呈並聯關係。梭內肌纖維的收縮成分位於纖維的兩端,而感受裝置位於其中間部,兩者呈串聯關係。因此,當梭外肌纖維收縮時,感受裝置所受的牽拉刺激將減少;而當梭內肌纖維收縮時,則感受裝置對牽拉刺激的敏感度增高。肌梭的傳入神經支配有兩類。I類傳入纖維直徑較粗(12-20μm),Ⅱ類傳入纖維直徑較細(4-12μm)。中樞有運動傳出支配梭外肌和梭內肌纖維,前者稱為α傳出纖維(直徑12-20μm),後者稱為γ傳出纖維(直徑2-6μm)。當γ傳出纖維活動加強時,梭內肌纖維收縮,可提高肌梭內感受裝置的敏感性,因此γ傳出纖維的活動對調節牽張反射具有重要作用。進一步研究指出,梭內肌纖維分兩類:一類其細胞核集中於中央部稱為核袋纖維(nuclear bag fiber),它接受γ1傳出纖維支配,並對快速牽拉較敏感,其傳入纖維主要屬I類;另一類其細胞核分散於整個纖維稱為核鏈纖維(nuclearchain fiber),它接受γ2傳出纖維支配,並對緩慢持續牽拉較敏感,其傳入纖維有I類和Ⅱ類(圖10-31)。
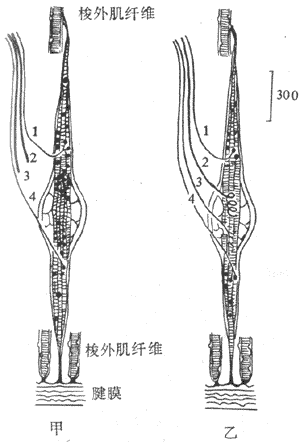
圖10-31 兩類梭內肌纖維示意圖
A:核袋纖維 B:核鏈纖維
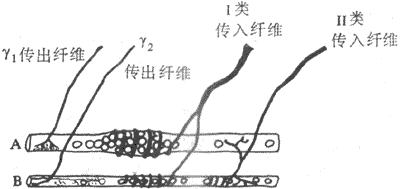
腱器官分布在肌腱膠原纖維之間的牽張感受裝置,由較細的I類纖維(直徑12μm)支配,末梢一般只有幾個分支。腱器官與梭外肌纖維呈串聯關係,其功能與肌梭功能不同,是感受骨肉張力變化的裝置。當梭外肌纖維發生等長收縮時,腱器官的傳入衝動發放頻率不變,肌梭的傳入衝動頻率減少;當肌肉受到被動牽拉時,腱器官和肌梭的傳入衝動發放頻率均增加。因此,腱器官是一種張力感受器,而肌梭是一種長度感受器。此外,腱器官的傳入衝動對同一肌肉的α運動神經元起牽拉抑制作用,而肌梭的傳入衝動對同一肌肉的α運動神經元起興奮作用。一般認為,當肌肉受到牽拉時,首先興奮肌梭的感受裝置發動牽張反射,引致受牽拉的肌肉收縮以對抗牽拉;當牽拉力量進一步加大時,則可興奮腱器官使牽張反射受抑制,以避免被牽拉的肌肉受到損傷。
(四)節間反射
脊動物在反射恢復的後期,可出現複雜的節間反射。例如,刺激動物腰背皮膚,可引致後肢發生一系列節奏性搔爬動作,稱為搔爬反射。搔爬反向依靠脊髓上下節段的協同活動,所以是節間反射的一種表現。
| 關於「生理學/脊髓對軀體運動的調節」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |