生理學/胃的分泌
| 醫學電子書 >> 《生理學》 >> 消化和吸收 >> 胃內消化 >> 胃的分泌 |
| 生理學 |
|
|
胃粘膜是一個複雜的分泌器官,含有三咱管狀外分泌腺和多種內分泌細胞。
胃的外分泌腺有:①賁門腺分布在胃與食管連接處的寬約1-4cm的環狀區內,為粘液腺,分泌粘液;②泌酸腺分布在佔全胃粘膜約2/3的胃底和胃體部。泌酸腺由三種細胞組成:壁細胞、主細胞和粘液頸細胞,它們分別分泌鹽酸、胃蛋白酶原和粘液;③幽門腺分布在幽門部,是分泌鹼性粘液的腺體。胃液是由這三種腺體和胃粘膜上皮細胞的分泌物構成的。
胃粘膜內至少含有6種內分泌細胞,如分泌胃泌素的G細胞、分泌生長抑素的D細胞和分泌組胺的肥大細胞等。
(一)胃液的性質、成分和作用
純淨的胃液是一種無色而呈酸性反應的液體,Pho 0.9-1.5。正常人每日分泌的胃液量約為1.5-2.5L。胃液的成分包括無機物如鹽酸、鈉和鉀的氯化物等,以及有機物如粘蛋白、消化酶等。與唾液相似,胃液的成分也隨分泌的速率而變化,當分泌率增加時,氫離子濃度升高,鈉離子濃度下降,但氯和鉀的濃度幾乎保持恆定(圖6-10)。
1.鹽酸 胃液中的鹽酸也稱胃酸,其含量通常以單位時間內分泌的鹽酸mmol表示,稱為鹽酸排出量。正常人空腹時鹽酸排出量(基礎酸排出量)約為0-5mmol/h。在食物或藥物(胃泌素或組胺)的刺激下,鹽酸排出量可進一步增加。正常人的鹽酸最大排出量可達20-25mmol/h。男性的酸分泌多於女性;鹽酸的排出量反映胃的分泌能力,它主要取決於壁細胞的數量(圖6-11),但也與壁細胞的功能狀態有關。
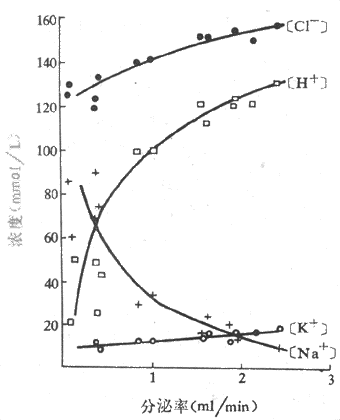
圖6-10 人胃液中電解質濃度與分泌率的關係胃液分泌是用組胺靜脈注射引起的
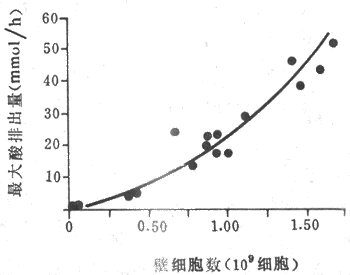
圖6-11 胃酸最大排出量與壁細胞數目的關係
由17個人胃的切除部分計算出最大酸排出量和壁細胞數目的關係。
由圖顯示每100萬個壁細胞可產酸約25mEp/h
胃液中H+的最大濃度可達150mmol/L,比血液中H+的濃度高三、四百萬倍,因此,壁細胞分泌H+是逆著巨大的濃度梯度進行的,需要消耗大量的能量,能量來源於氧代謝。
泌酸所需的H+來自壁細胞漿內的水。水解離產生H+和OH-,任借存在於壁細胞上分泌小管膜上的H+、K+-ATP酶的作用,H+被主動地轉運入小管腔內。
壁細胞分泌小管膜上的H+、K+-ATP酶又稱質子泵(proton pump)或稱酸泵。H+-K+交換是壁細胞質子泵區別於體內任何其它細胞上的質子泵的顯著特徵。H+、K+-ATP酶每催化一分子的ATP分解為ADP和磷酸所釋放的能量,可驅動一個H+從壁細胞漿進入分泌小管腔和一個K+從小管腔進入細胞漿。H+的分泌必須在分泌小管內存在足夠濃度的K+的條件下才能進行。
年來,選拔性干擾胃壁細胞的H+、K+-ATP酶的藥物已被用來有效地抑制胃酸分泌,成為一代新型的抗潰瘍藥物。
已知壁細胞內含有豐富的碳酸酐酶,在它的催化下,由細胞代謝產生的CO2和由血漿中攝取的CO2可迅速地水合而形成H2CO3,H2CO3隨即又解離為H+和HCO3。這樣,在H+分泌後,留在細胞內的OH-便和由
H2CO3解離的H+結合而被中和,壁細胞內將不致因OH-的蓄積而使pH升高。由H2CO3產生的HCO3則在壁細胞的底側膜,與CI-並換而進入血液。因此,餐後與大量胃酸分泌的同時,血和尿的pH往往升高而出現「餐後鹼潮」。與HCO3交換而進入壁細胞內的CI-則通過分泌小管膜上特異性的CI-通道進入小管腔,與H+形成HCi (圖6-12)。
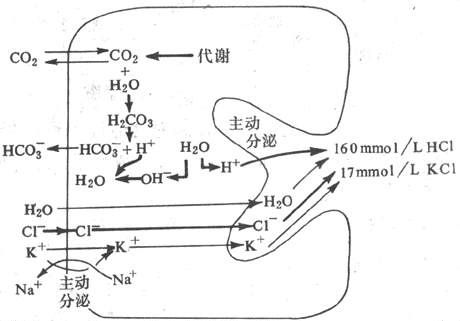
圖6-12 壁細胞分泌鹽酸的一種假設
胃內的鹽酸有許多作用,它可殺死隨食物進入胃內的細菌,因而對維持胃和小腸內的無菌狀態具有重要意義。鹽酸還能激活胃蛋白酶原,使之轉變為有活性的胃蛋白酶,鹽酸並為胃蛋白酶作用提供了必要的酸性環境。鹽酸進入小腸後,可以引起促胰液素的釋放,從而促進胰液、膽汁和小腸液的分泌。鹽酸所造成的酸性環境,還有助於小腸對鐵和鈣的吸收。但若鹽酸分泌過多,也會對人體產生不利影響,。一般認為,過高的胃酸對胃和十二指腸粘膜有侵蝕作用,因而潰瘍病發病的重要原因之一。
2.胃蛋白酶原 胃蛋白酶原是由主細胞合成的,並以不具有活性的酶原顆粒形式貯存在細胞內。當細胞內充滿酶原顆粒時,它對新的酶原的全盛2產生負反饋作用。持續的刺激可使主細胞內的顆粒歷釋放而完全消失,便分泌仍繼續進入,說明酶原也可以不經過顆粒的形式直接釋放出來。
分泌入胃腔內的胃蛋白酶原在胃酸的作用下,從分子中分離出一個小分子的多肽,轉變為具有活性的胃蛋白酶。已激活的胃蛋白酶對胃蛋白酶原也有激活作用。
胃蛋白酶能水解食物中的蛋白質,它主要作用於蛋白質及多肽分子中含苯丙氨酸或酪氨酸的肽鍵上,其主要分解產物是腖,產生多肽或胺基酸較少。胃蛋白酶只有在酸性較強的環境中才能發揮作用,其最知pH為2。隨著pH的升高,胃蛋白酶的活性即降低,當pH升至6以上時,此酶即發生不可逆的變性。
3.粘液和碳酸氫鹽 胃的粘液是由表面上皮細胞、泌酸腺的粘液頸細胞,賁門腺和幽門腺共同分泌的,其主要成分為糖蛋白。糖蛋白是由4個亞單位通過二硫鍵連接形成的(圖6-13)。由於糖蛋白的結構特點,粘液具有較高的粘滯性和形成凝膠的特性。在正常人,粘液覆蓋在胃粘膜的表面,形成一個厚約500μm的凝膠層,它具有潤滑作用,可減少粗糙的食物對胃粘膜的機械性損傷。
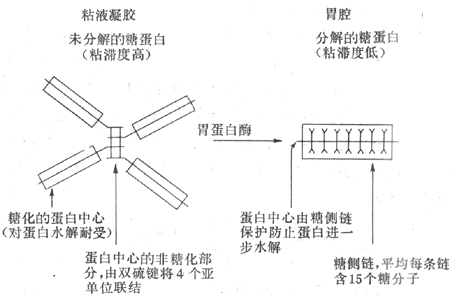
圖6-13 胃粘液糖蛋白結構的示意圖
胃內HCO3主要是由胃粘膜的非泌酸細胞分泌的,僅有少量的HCO3是從組織間液滲入胃內的。基礎狀態下,胃HCO3分泌的速率僅為H+分泌速率的5%。進食時其分泌速率的增加通常是與H+分泌速率的變化平行的。由於H+和HCO3在分泌速率和濃度上的巨大差距,分泌的HCO3對辦內pH顯然不會有多大影響。
長期以來人們一直在思索:胃粘膜處於高酸和胃蛋白酶的環境中,為什麼不被消化?近年來「粘液-碳酸氫鹽屏障」概念的提出,至少部分地回答了這個問題。這主要是因為。胃粘液的粘稠度約為水的30-260倍,H+和HCO3電漿在粘液層內的擴散速度明顯減慢,因此,在胃腔同內的H+向粘液凝膠深層彌散過程中,它不斷地與從粘液層下面的上皮細胞分泌並向表面擴散的HCO3遭遇,兩種離子在粘液層內發生中和。用pH測量電極測得,在胃粘液層存在一個pH梯度,粘液層靠近胃腔面的一側呈酸性,pH為7左右(圖6-14)。因此,由沾液和碳酸氫鹽共同構築的粘液-碳酸氫鹽屏障。能有效地阻擋H+的逆向彌散,保護了胃粘液免受H+的假侵蝕;粘液深層的中性pH環境還使胃蛋白酶喪失了分解蛋白質的作用。
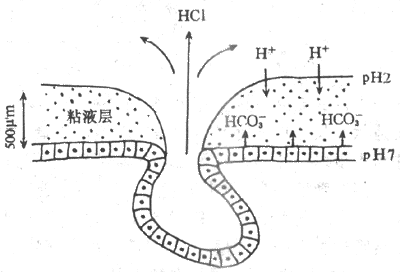
圖6-14 胃粘液-碳酸氫鹽屏障模式圖
正常情況下,胃粘液凝膠層臨近胃腔一側的糖蛋白容易受到胃蛋白酶的作用而水解為4個亞單位,這樣,粘液便從凝膠狀態變為溶膠狀態而進入胃液。但一般來講,水解的速度與粘膜上皮細胞分泌液的速度相等,這種粘液分泌與降解裼動態平衡,保持了粘液屏障功能的完整性和連續性。
4.內因子泌酸腺的壁細胞除分泌鹽酸外,還分泌一種分子量在50000-60000之間的糖蛋白,稱為內因子。內因子可與進入胃內的維生素B12結合而促進其吸收。
(二)胃液分泌的調節
胃液分泌受許多因素的影響,其中有的起興奮性作用,有的則起抑制性作用。進食是胃液分泌的自然刺激物,它通過神經和體液因素調節胃液的分泌。
1.刺激胃酸分泌的內源性物質
(1)乙醯膽鹼:大部分支配胃的副交感神經節後纖維末稍釋放乙醯膽鹼。乙醯膽鹼直接作用於壁細胞膜上的膽鹼能受體,引起鹽酸分泌增加。乙醯膽鹼的作用可被膽鹼能受體阻斷劑(如阿托品)阻斷。
(2)胃泌素:胃泌素主要由胃竇粘膜內的G細胞分泌。十二指腸和空腸上段粘膜內也有少量G細胞。胃泌素釋放後主要通過血液循環作用於壁細胞,刺激其分泌鹽酸。
胃泌素以多種分子形式存在於體內,其主要的分子形式有兩種:大胃泌素(G-34)和小胃泌素(G-17)。胃竇粘膜內的胃泌素主要是G-17,十二指腸粘膜中有G-17和G-34約各佔一半。從生物效應來看,G-17刺激胃分泌的作用要比G-34強5-6倍,但G-34在體內被清除的速度很慢,它攔衰期約為50min,而G-17通常只有6min。
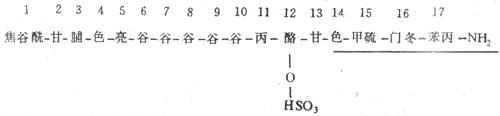
圖6-15 胃泌素的胺基酸序列
劃線的部分是其活性片段
人的小胃泌素的胺基酸序列如圖6-15,其C端正的4個胺基酸是胃泌素的最小活性片段,因此,用人工合成的四肽或五肽胃泌素是具有天然胃泌全部作用的人工製品。
(三)組胺:胃的泌酸區
粘膜內含有大量的組胺。產生組胺的細胞是存在於固有膜中的肥大細胞。正常情況下,胃粘膜恆定地釋放少量組胺,通過局部彌散到達鄰近的壁細胞,刺激其分泌。壁細胞上的組胺受體為Ⅱ型受體(H2受體),用甲氰咪呱(cimetidine)及其相類似的藥物可以阻斷組胺與壁細胞的結合,從而減少胃酸分泌。
以上三種內源性分泌物,一方面可通過各自壁細胞上的特異性受體,獨立地發揮刺激胃酸分泌的作用(圖6-16);另一方面,三者又相互影響,表現為當以上三個因素中的兩個因素同時作用時,胃酸的分泌反應往往比這兩個因素單獨作用的總和要大,這種現象在生理學上稱為加強作用(potentiation)。在整體內,促分泌物之間的相互加強作用是經常存在的,因此,用任何一咱促分泌物的阻斷劑,如用甲氰咪呱時,它不僅抑制了壁細胞對組胺的反應,同時也會由於去除了組胺的作用的背景,使壁細胞對胃泌素和乙醯膽鹼的反應也有所降低。
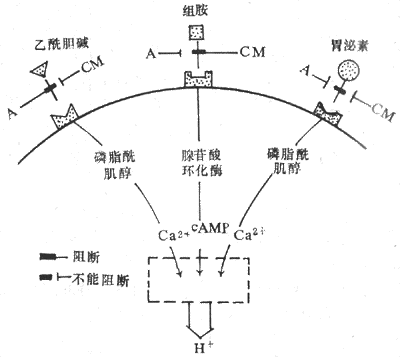
圖6-16 三種刺激胃酸分泌的內源性物質的作用及相互關係
A:阿托品 CM:甲氰咪呱
2.消化期的胃液分泌 進食後胃液分泌的機制,一般按接受食物刺激的部位,分成三個時期來分析,即頭期、胃期和腸期。但必須注意,三個時期的劃分是人為的,只是為了便於敘述,實際上,這三個時期幾乎是同時開始的、相互重疊的。
(1)頭期胃液分泌:頭期的胃液分泌是由進食動作引起的,因其傳入衝動均來自頭部感受器(眼、耳、口腔、咽、食管等)。因而稱為頭期。
頭期胃液分泌的機制曾用慢性方法作了較詳細的分析,即用事先旅行過食管切斷術並具有胃瘺的狗進行假飼(sham feeding);當食物經過口腔進入食管後,隨即從食管的切品流出體外,食物並未進入胃內,但卻引起胃液分泌。進一步分析確定,由進食動作所引起的胃液分泌,包括條件反射性和非條例反射性兩種分泌。前者是由和食物有關的形象、氣味、聲音等刺激了視、嗅、聽等感受器而引起的;後者則當咀嚼和吞咽食物時,刺激了口腔和咽喉等外的化學和機械感受器而引起的。這些反射的傳入途徑和由進食引起的唾液分泌的傳入途徑相同,反射中樞包括延髓、下丘腦、邊緣和大腦皮層等。迷走神經是這些反射共同的傳出神經。當切斷支配胃的迷走神經後,假飼就不再引起胃液分泌。
迷走神經興奮後,除了通過其末稍釋放乙醯膽鹼,直接引起腺體細胞分泌外,迷走神經衝動還可引起胃竇粘膜內的G細胞釋放胃泌素,後者經過血液循環刺激胃腺頒發(圖6-17)。由此可見,頭期的胃注分泌並不是純神經反射性的,而是一種神經-體液性的調節。
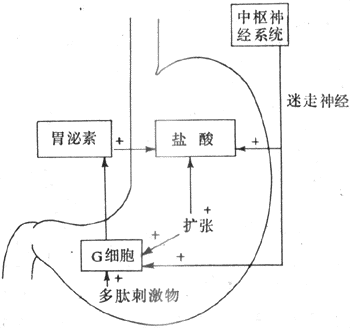
圖6-17 引起胃酸分泌的機制
引起胃泌素釋放的迷走神經纖維被認為是非膽鹼能的,因為阿托品不僅不能阻斷,反而使假飼引起的胃泌素釋放反應增加。目前對這一現象的解釋是,迷走神經中既存在興奮胃泌素釋放的纖維,也存在抑制胃泌素釋放的纖維,前者的中介物可能是一種肽類物質,而抑制性纖維則是通過乙醯膽鹼能起作用的。阿托品由於阻斷了抑制性纖維的作用,因而使胃泌素的釋放有所增加。
頭期胃液分泌的量和酸度都很高,而胃蛋白酶的含量則尤其高。在人體觀察的資料表明,頭期胃液分泌的大小與食慾有很大的關係。
(2)胃期胃液分泌:食物入胃後,對胃產生機械性和化學性刺激,繼續引起胃液分泌,其主要途徑為:①擴張刺激胃底、胃體部的感受器,通過迷走、迷走神經經長反射和壁內神經叢的短反射,引起胃腺分泌;②擴張刺激胃幽門部,通過壁內神經叢,作用於G細胞,引起胃泌素的釋放;③食物的化學成分直接作用於G細胞,引起胃泌素的釋放。
刺激G細胞釋放胃泌素的主要食物化學成分是蛋白質的消化產物,其中包括肽類和胺基酸。G細胞為開放型胃腸內分泌細胞,頂端有絨毛樣突起伸入胃腔,可以直接感受胃腔內化學物質的作用。用放射免疫方法測定血漿中胃泌素濃度,正常人空腹時約為30-120pg/ml,在進食蛋白質食物後,血漿胃泌素可升高到50-200pg/ml,在食後2-3小時逐漸恢復至進食前水平。糖類和脂肪類食物不是胃泌素釋放的強刺激物。
胃酸分泌的胃液酸度也很高,但胃蛋白酶含量卻比頭期分泌的胃液為弱。
(3)腸期胃液分泌:將食糜、內的提取液、蛋白腖液由瘺管直接注入十二指腸內,也可引起胃液分泌的輕度增加,說明當食物離開胃進入小腸後,還有繼續刺激胃液分泌的作用。機械擴張游離的空腸袢,胃液分泌也增加。
在切斷支配胃的外來神經後,食物對小腸的作用仍可引起胃液分泌,提示腸期胃液分泌的機制中,神經反射的作用不大,它主要通過體液調節機制,即當食物與小腸粘膜接觸時,有一種或幾種激素從小腸粘膜釋放出來,通過血液循環作用於胃。已知人的十二指腸粘膜中含有較多的胃泌素;用放射免疫方法測得,在切除了胃竇的病人,進食後血漿胃泌素的濃度仍有升高,說明進食後可引起十二指腸釋放胃泌素,它可能是腸期胃泌分泌的體液因素之一。有人認為,在食糜作用下,小腸粘膜還可能釋放腸泌酸素的激素,刺激胃酸分泌。此外,由小腸吸收的胺基酸也可能參與腸期的胃液分泌,因為靜脈注射混合胺基酸也可引起胃酸分泌。
腸期胃液分泌的量不大,大約佔進食後胃液分泌總量的1/10,這可能與食物在小腸內同時還產生許多對胃液起抑制性作用的調節(見後文)有關。
3.胃液分泌的抑制性調節上已述在進食過程中興奮胃液分泌的機制,而正常消化期的胃液分泌還受到各種抑制性因素的調節,實際表現的胃液分泌正是興奮和抑制性因素共同作用的結果。在消化期同內,抑制胃液分泌的因素除精神、情緒因素外,主要在鹽酸、脂肪和高張溶液三種。
鹽酸是胃腺活動的產物,但它對胃腺的活動又具有抑制性作用,因此是胃酸分泌的一種負反饋的調節機制。
當胃竇的pH降到1.2-1.5時,便可能對胃液分泌的產生抑制作用。這種抑制作用的機制可能是鹽酸直接抑制了胃竇粘膜中的G細胞,減少胃泌素釋放的結果。惡性貧血的病人胃酸分泌很低,他們血漿中胃泌素的濃度卻比正常人高20-30倍,如向這種病人胃內注以鹽酸,使胃內酸化,血漿胃泌素的濃度即下降,這進一步說明,胃內容物的酸度對胃泌素的釋放,以及進而影響胃液分泌具有重要作用。
近年來一些實驗資料提示,鹽酸在胃內還可能通過引起胃粘膜釋放一種抑制性因素——生長抑素,轉而抑制胃泌素和胃液的分泌。
當十二接指腸內的pH降到2.5以下時,對胃酸分泌也有抑制作用,但其作用機制目前尚未完全闡明。已知酸作用於小腸粘膜可引起促胰液素釋放,後者對胃泌素引起的酸分泌具有明顯的抑制作用,因此,促胰液素很可能是十二指腸酸化抑制胃分泌的一種抑制物。此外,十二指腸球部在鹽酸刺激下,也可能釋放出一種抑制胃分泌的肽類激素——球抑胃素,但球抑胃素結構尚未最後確定。
脂肪是抑制胃液分泌的一個重要因素。脂肪及其消化產物抑制胃分泌的作用發生在脂肪進入十二指腸後,而不是在胃中。早在30年代,我國生理學家林可勝就發現,從小腸粘膜中可提取出一種物質,當由靜脈注射後,保使胃液分泌的量、酸度和消化力減低,並抑制胃運動。這個物質被認為是脂肪在小腸內抑制胃分泌的體液因素,而可能是幾種具有此種作用的激素的總稱。小腸粘膜中存在的抑胃肽、神經降壓素等多種激素,都具有類似腸抑素的特性。
十二指腸內高張溶液對胃分泌的抑制作用可能通過兩種途徑來實現,即汽船活小腸內滲透壓感受器,通過腸-胃反射引起胃酸分泌的抑制,以及通過刺激小腸粘膜釋放一種或幾種抑制性激素而抑制胃液分泌。後一機制尚未闡明。
在胃的粘膜和肌層中,存在大量的前列腺素(詳見內分泌章)。迷走神經興奮和胃泌素都可引起前列腺素釋放的增加。前列腺素對進食、組胺和胃泌素等引起的胃液分泌有明顯的抑制作用。它可能是胃液分泌的負反饋抑制物。前列腺素還能減少胃粘膜血流,但它抑制胃分泌的作用並非繼發於血流的改變。
| 關於「生理學/胃的分泌」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |