生物化學與分子生物學/糖異生
| 醫學電子書 >> 《生物化學與分子生物學》 >> 糖代謝 >> 糖異生 |
| 生物化學與分子生物學 |
|
|
非糖物質轉變為葡萄糖或糖原的過程稱為糖異生(gluconeogenesis)。非糖物質主要有生糖胺基酸(甘、丙、蘇、絲、天冬、谷、半胱、脯、精、組等)、有機酸(乳酸、丙酮酸及三羧酸循環中各種羧酸等)和甘油等。不同物質轉變為糖的速度不同。
進行糖異生的器官,首推肝臟,長期飢餓和酸中毒時腎臟中的糖異生作用大大加強,相當於同重量的肝組織的作用。
目錄 |
一、糖異生的途徑
糖異生的途徑基本上是糖酵解或糖有氧氧化的逆過程,糖酵解通路中大多數的酶促反應是可逆的,但是糖酵解途徑中己糖激酶、磷酸果糖激酶和丙酮酸激酶三個限速酶催化的三個反應過程,都有相當大的能量變化,因為己糖激酶(包括葡萄糖激酶)和磷酸果糖激酶所催化的反應都要消耗ATP而釋放能量,丙酮酸激酶催化的反應使磷酸烯醇式丙酮酸轉移其能量及磷酸基生成ATP,這些反應的逆過程就需要吸收相等量的能量,因而構成「能障」,為越過障礙,實現糖異生,可以由另外不同的酶來催化逆行過程,而繞過各自能障,這種由不同的酶催化的單向反應,造成兩個作用物互變的循環稱為作用物循環或底物循環。
(一)由丙酮酸激酶催化的逆反應是由兩步反應來完成的。
首先由丙酮酸羧化酶催化,將丙酮酸轉變為草醯乙酸,然後再由磷酸烯醇式丙酮酸羧激酶催化,由草醯乙酸生成磷酸烯醇式丙酮酸。
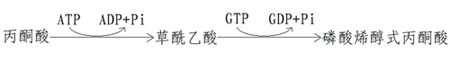
這個過程中消耗兩個高能鍵(一個來自ATP,另一個來自GTP),而由磷酸烯醇式丙酮酸分解為丙酮酸只生成1個ATP。
由於丙酮酸羧化酶僅存在於粒線體內,胞液中的丙酮酸必須進入粒線體,才能羧化生成草醯乙酸,而磷酸烯醇式丙酮酸羧激酶在粒線體和胞液中都存在,因此草醯乙酸可在粒線體中直接轉變為磷酸烯醇式丙酮酸再進入胞液中,也可在胞液中被轉變為磷酸烯醇式丙酮酸。但是,草醯乙酸不能通過粒線體膜,其進入胞液可通過兩種方式將其轉運:一種是經蘋果酸脫氫酶作用,將其還原成蘋果酸,然後通過粒線體膜進入胞液,再由胞液中NAD+-蘋果酸脫氫酶將蘋果酸脫氫氧化為草醯乙酸而進入糖異生反應途徑,由此可見,以蘋果酸代替草醯乙酸透過粒線體膜不僅解決了糖異生所需要的碳單位,同時又從粒線體內帶出一對氫,以NADH+H+形成使1,3-二磷酸甘油酸生成3磷酸甘油醛,從而保證了糖異生順利進行。另一種方式是經穀草轉氨酶的作用,生成天門冬氨酸後再逸出粒線體,進入胞液中的天門冬氨酸再經胞液中穀草轉氨酶催化而恢復生成草醯乙醯。有實驗表明,以丙酮酸或能轉變為丙酮酸的某些成糖胺基酸作為原料成糖時,以蘋果酸通過粒線體方式進行糖異生,而乳糖進行糖異生反應時,它在胞液中變成丙酮酸時已脫氫生成NADH+H+,可供利用,故常在粒線體內生成草醯乙酸後,再變成天門冬氨酸而出粒線體內膜進入胞漿(圖4-10)。
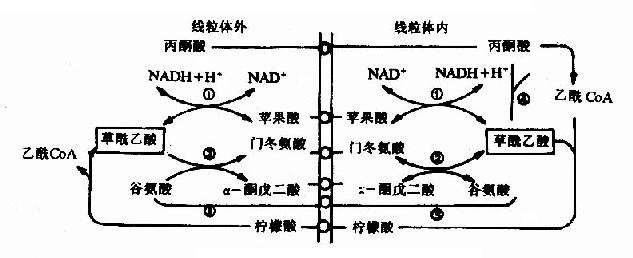
圖4-10 草酸乙酸逸出粒線體方式
①蘋果酸脫氫酶 ②穀草轉氨酶 ③檸檬合成酶 ④丙酮酸羧化酶 ⑤ATP-檸檬裂酸酶合
(二)由己糖激酶和磷酸果糖激酶催化的兩個反應的逆行過程
由兩個特異的磷酸酶水解己糖磷酸酯鍵完成,催化G-6-P水解生成葡萄糖的酶為葡萄糖-6-磷酸酶(glucose-6-phosphatase);催化1,6-二磷酸果糖水解生成F-6-P的酶是果糖二磷酸酶(fructose diphosphatase)。
除上述幾步反應以外,糖異生反應就是糖酵解途徑的逆反應過程。因此,糖異生可總結為:
2丙酮酸+4ATP+2GTP+2NADH+2H+ +6H2O→葡萄糖+2NAD++4ADP+2GDP+6Pi+6H+
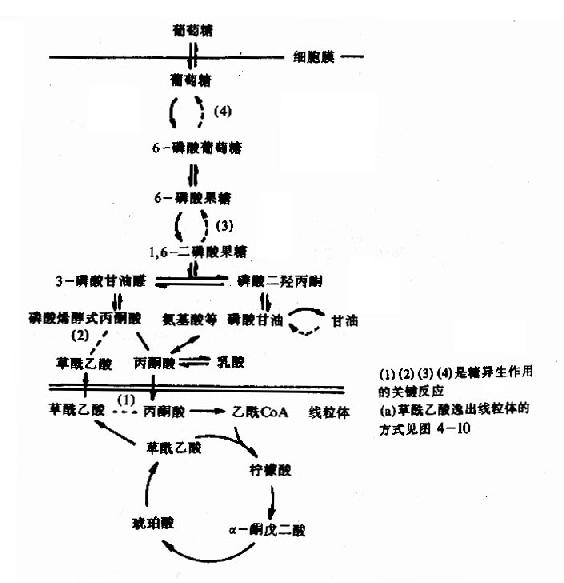
圖4-11 肝與腎皮質中糖氧化與糖異生的通路
現將肝臟和腎皮質中糖的氧化與糖異生作用過程總結如圖4?1,糖異生作用的三種主要原料有乳酸、甘油和胺基酸等,乳酸在乳酸脫氫酶作用下轉變為丙酮酸,經前述羧化支路成糖;甘油被磷酸化生成磷酸甘油後,氧化成磷酸二羥丙酮,再循糖酵解逆行過程合成糖;胺基酸則通過多種渠道成為糖酵解或糖有氧氧化過程中的中間產物,然後生成糖;三羧酸循環中的各種羧酸則可轉變為草醯乙酸,然後生成糖。
二、糖異生的生理意義
(一)糖異生作用的主要生理意義
是保證在飢餓情況下,血糖濃度的相對恆定。
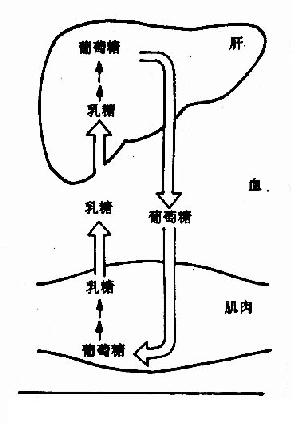
圖4-12 Cori循環
血糖的正常濃度為3.89?.11mmol/L,即使禁食數周,血糖濃度仍可保持在3.40mmol/L左右,這對保證某些主要依賴葡萄糖供能的組織的功能具有重要意義,停食一夜(8-10小時)處於安靜狀態的正常人每日體內葡萄糖利用,腦約125g,肌肉(休息狀態)約50g,血細胞等約50g,僅這幾種組織消耗糖量達225g,體內貯存可供利用的糖約150g,貯糖量最多的肌糖原僅供本身氧化供能,若只用肝糖原的貯存量來維持血糖濃度最多不超過12小時,由此可見糖異生的重要性。
(二)糖異生作用與乳酸的作用密切關係
在激烈運動時,肌肉糖酵解生成大量乳酸,後者經血液運到肝臟可再合成肝糖原和葡萄糖,因而使不能直接產生葡萄糖的肌糖原間接變成血糖,並且有利於回收乳酸分子中的能量,更新肌糖原,防止乳酸酸中毒的發生。(圖4-12)
(三)協助胺基酸代謝
實驗證實進食蛋白質後,肝中糖原含量增加;禁食晚期、糖尿病或皮質醇過多時,由於組織蛋白質分解,血漿胺基酸增多,糖的異生作用增強,因而胺基酸成糖可能是胺基酸代謝的主要途徑。
(四)促進腎小管泌氨的作用
長期禁食後腎臟的糖異生可以明顯增加,發生這一變化的原因可能是飢餓造成的代謝性酸中毒,體液pH降低可以促進腎小管中磷酸烯醇式丙酮酸羧激酶的合成,使成糖作用增加,當腎臟中α酮戊二酸經草醯乙酸而加速成糖後,可因α-酮戊二酸的減少而促進谷氨醯胺脫氨成谷氨酸以及谷氨酸的脫氨,腎小管細胞將NH3分泌入管腔中,與原尿中H+結合,降低原尿H+的濃度,有利於排氫保納作用的進行,對於防止酸中毒有重要作用。
三、糖異生的調節
糖異生的限速酶主要有以下4個酶:丙酮酸羧化酶、磷酸烯醇式丙酮酸羧激酶、果糖二磷酸酶和葡萄糖磷酸酶。
(一)激素對糖異生的調節
激素調節糖異生作用對維持機體的恆穩狀態十分重要,激素對糖異生調節實質是調節糖異生和糖酵解這兩個途徑的調節酶以及控制供應肝臟的脂肪酸,更大量的脂肪酸的獲得使肝臟氧化更多的脂肪酸,也就促進葡萄糖合成,胰高血糖素促進脂肪組織分解脂肪,增加血漿脂肪酸,所以促進糖異生;而胰島素的作用則正相反。胰高血糖素和胰島素都可通過影響肝臟酶的磷酸化修飾狀態來調節糖異生作用,胰高血糖素激活腺苷酸環化酶以產生cAMP,也就激活cAMP依賴的蛋白激酶,後者磷酸化丙酮酸激酶而使之抑制,這一酵解途徑上的調節酶受抑制就刺激糖異生途徑,因為阻止磷酸烯醇式丙酮酸向丙酮酸轉變。胰高血糖素降低2,6-二磷酸果糖在肝臟的濃度而促進1,6-二磷酸果糖轉變為6磷酸果糖,這是由於2,6-二磷酸果糖是果糖二磷酸酶的別位抑制物,又是6磷酸果糖激酶的別位激活物,胰高血糖素能通過cAMP促進雙功能酶(6磷酸果糖激酶2/果糖2,6-二磷酸酶)磷酸化。這個酶經磷酸化後就滅活激酶部位卻活化磷酸酶部位,因而2,6-二磷酸果糖生成減少而被水解為6磷酸果糖增多。這種由胰高血糖素引致的2,6-二磷酸果糖下降的結果是6磷酸果糖激酶1活性下降,果糖二磷酸酶活性增高,果糖二磷酸轉變為6磷酸果糖增多,有利糖異生,而胰島素的作用正相反(圖4-13)。
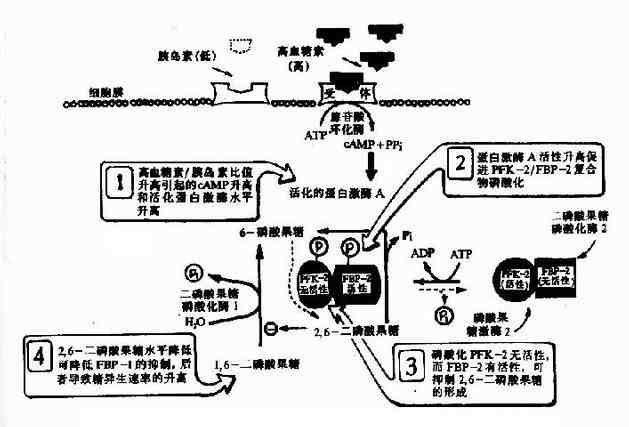
圖4-13 高血糖素水平升高對肝細胞外2,6-二磷酸果糖濃度的影響
除上述胰高血糖素和胰島素對糖異生和糖酵解的短快調節,它們還分別誘導或阻遏糖異生和糖酵解的調節酶,胰高血糖素/胰島素比例高誘導大量磷酸烯醇式丙酮酸羧激酶,果糖6-磷酸酶等糖異生酶合成而阻遏葡萄糖激酶和丙酮酸激酶的合成。
(二)代謝物對糖異生的調節
1.糖異生原料的濃度對糖異生作用的調節血漿中甘油、乳酸和胺基酸濃度增加時,使糖的異生作用增強。例如飢餓情況下,脂肪動員增加,組織蛋白質分解加強,血漿甘油和胺基酸增高;激烈運動時,血乳酸含量劇增,都可促進糖異生作用。
2.乙醯輔酶A濃度對糖異生的影響乙醯輔酶A決定了丙酮酸代謝的方向,脂肪酸氧化分解產生大量的乙醯輔酶A可以抑制丙酮酸脫氫酶系,使丙酮酸大量蓄積,為糖異生提供原料,同時又可激活丙酮酸羧化酶,加速丙酮酸生成草醯乙酸,使糖異生作用增強。
此外乙醯CoA與草醯乙酸縮合生成檸檬酸由粒線體內透出而進入細胞液中,可以抑制磷酸果糖激酶,使果糖二磷酸酶活性升高,促進糖異生。
| 關於「生物化學與分子生物學/糖異生」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |