生理學/紅細胞生理
| 醫學電子書 >> 《生理學》 >> 血液 >> 血細胞及其功能 >> 紅細胞生理 |
| 生理學 |
|
|
1.紅細胞的數量、形態和功能 紅細胞(erythuocyte)是血液中數量最多的一種血細胞,正常男性每微升血液中平均約500萬個(5.0×1012/L),女性較少,平均約420萬個(4.2×1012/L).紅細胞含有血紅蛋白,因而使血液呈紅色.紅細胞在血液的氣體運輸中有極重要的作用.在血液中由紅細胞運輸的氧約為溶解於血漿的70倍;在紅細胞參與下,血漿運輸二氧化碳的能力約為直接溶解於血漿的18倍(詳見第五章第三節).正常紅細胞呈雙凹圓碟形,平均直徑約8μm,周邊稍厚.這種細胞開頭的表面積與體積之比,較球形時為大,因而氣體可通過的面積也較大;由細胞中心到大部分表面的距離較短,因此氣體進出紅細胞的擴散距離也較短.這種形狀也有利於紅細胞的可塑性變形.紅細胞在全身血管中循環運行,常要擠過口徑比它小的毛細血管和血竇間隙,這時紅細胞將發生捲曲變形,在通過後又恢復原狀,這種變形稱為塑性變形.表面積與體積的比值愈大,變形能力愈大,故雙凹圓碟形紅細胞的變形能力遠大於異常情況下可能出現的球形紅細胞.紅細胞保持雙凹圓碟形需要消耗能量。
紅細胞膜是以脂質雙分子層為骨架的半透膜。氧和二氧化碳等脂溶性氣體可以自由通過,尿素也可以自由透入。在電解質中,負離子(如CI-、HCO3-)一般較易通過紅細胞膜,而正離子卻很難通過。紅細胞內Na+ 濃度遠低於細胞外,而細胞內K+濃度遠高於細胞外,這種細胞內外的Na+、K+濃度差主要是依靠細胞膜上Na+泵的活動來維持的。低溫貯存較久的血液,血漿內K+濃度升高,就是由於低溫下代謝幾乎停止,Na+泵不能活動的緣故。
紅細胞結合和攜帶氧的過程並不消耗能量,血紅蛋白中的Fe2+也不被氧化,若Fe2+被氧化成Fe3+成為高鐵血紅蛋白,即失去攜氧能力。紅細胞消耗葡萄糖,主要是通過糖酵解和磷酸戊糖旁路,所產生的能量(以結合於ATP的形式)主要是用於供應細胞膜上Na+泵的活動,用於保持低鐵血紅蛋白不致被氧化,也用於保持紅細胞膜的完整性和細胞的雙凹圓碟形。
2.紅細胞比容 紅細胞在血液中所佔的容積百分比,稱為紅細胞比容(hematocritvalue),可以用分血計(hematocrit)來測定。通常是將一定量的血液與抗凝劑混勻,置於用直徑2.5mm的平底玻璃管製成的分血計中,以每分鐘3000轉的速度離心半小時,使血細胞下沉壓緊,即可測出紅細胞比容。正常成年人的紅細胞比容,男性為40%-50%,女性為37%-48%。但這是從手臂等處淺靜脈抽血測定的數值,並且這時在壓緊的紅細胞之間有很少量血漿;同時,全身各類血管中,血液的紅細胞比容值也不盡相同。
3.正常紅細胞生成所必需的原料和其它因素 在幼紅細胞的發育成熟過程中,細胞核的存在對於細胞分裂和合成血紅蛋白有著重要的作用。在這些階段,合成細胞核的主要構成物質—DNA必須有維生素B12和葉酸作為輔酶。
維生素B12是含鈷的有機化合物,多存在於動物性食品中。機體對維生素B12的吸收必須要有內因子(intrinsic factor)和R結合蛋白(Rprotein)參與。內因子是由胃腺的壁細胞所分泌的一種糖蛋白,分子量在50000-60000之間,而R(rapid)蛋白是一種電泳速度很快的血漿蛋白。在酸性的胃液中,維生素B12主要與R蛋白結合,到了小腸上段處胰蛋白酶將這種結合斷裂,維生素B12轉而與內因子結合。內因子有兩個活性部位,一個部位可與維生素B12結合,另一個部位則可與迴腸上皮細胞膜上的特異受體結合。在正常情況下,內因子-B12複合物在小腸上段可保護維生素B12不受小腸內蛋白水解酶的破壞。當複合物運行至迴腸段,便與迴腸粘膜受體結合而被吸收進入門脈系統血流,一部分貯存在肝,一部分又與運輸維生素B12的轉鈷蛋白Ⅱ(transcobalamineⅡ)結合,沿血液輸送到造血組織,參與紅細胞生成過程。當胃的大部分被切除或胃腺細胞受損傷,機體缺乏內因子,或體內產生抗內因子的抗體時,即可發生維生素B12吸收障礙,影響幼紅細胞的分裂和血紅蛋白合成,出現巨幼紅細胞性貧血,即大細胞性貧血。
葉酸是以蝶醯單谷氨酸的形式吸收的。吸收之後,在雙氫葉酸還原酶的催化下,形成四氫葉酸。存在於血漿中的葉酸幾乎全是四氫葉酸的單谷氨酸鹽。但進入組織細胞後,又通過酶促作用,再轉變為多谷氨酸鹽,才具有活性。葉酸缺乏時也引起與維生素B12缺乏時相似的巨幼紅細胞性貧血。只是在維生素B12缺乏時,還可伴有神經系統和消化道症状。
合成血紅蛋白還必須有鐵作為原料,每亳升紅細胞需要1mg鐵,每天需要20-25mg鐵用於紅細胞生成,但人每天只需從食物中吸收1mg(約5%)以補充排泄的鐵,其餘95%均來自人體鐵的再利用。機體貯存的鐵主要來自於破壞了的紅細胞。衰老的紅細胞被巨噬細胞吞噬後,血紅蛋白被消化而釋出血紅素中的Fe2+。這樣釋出的鐵即與鐵蛋白(ferritin)結合,此時的鐵為Fe3+,聚集成鐵黃素顆粒而沉澱於巨噬細胞內。血漿中有一種運鐵蛋白(transferrin),可以來往運行於巨噬細胞與幼紅細胞之間,以運送鐵。貯存於鐵蛋白中的Fe3+,先還原成Fe2+再脫離鐵蛋白,而後與運鐵蛋白結合。每分子運鐵蛋白可以運送兩個Fe2+,運送到幼紅細胞後,又可反覆作第二次運輸。此外,還可以通過巨噬細胞與紅母細胞直接接觸,以提供合成血紅蛋白所需的鐵。由於慢性出血等原因,體內貯存的鐵減少,或造血功能增強而供鐵不夠,均可引起小細胞性貧血,這主要是合成血紅蛋白不足。此外,紅細胞生成還需要胺基酸和蛋白質、維生素B6、B2、C、E,微量元素銅、錳、鈷和鋅等。
4.紅細胞生成的調節 每個成年人體內約有25×1012個紅細胞,每24小時便有0.8%的紅細胞進行更新,也就是說每分鐘約有160×106個紅細胞生成;當機體有需要時,如失血或某些疾病使紅細胞壽命縮短時,紅細胞的生成率還能在正常基礎上增加數倍。目前已經證明有兩種調節因子分別調製著兩個不同發育階段紅系祖細胞的生長。一種是早期的紅系祖細胞,稱為爆式紅系集落形成單位(burst forming unit-erythroid,BFU-E),這是因為它們在體外培養中能形成很大的細胞集落,組成集落的細胞散布成物體爆炸的形狀,這種早期祖細胞的生長和在體外形成集落都依賴於一種稱為爆式促進因子(burst promoting activitor,BPA)的刺激作用。BPA是一類分子量為25000-40000的糖蛋白,以早期紅系祖細胞BFU-E為作用的靶細胞,可能是促進更多的BFU-E從細胞周期中的靜息狀態(G0期)進入DNA合成期(S期),因而使早期祖細胞加強增殖活動。另一種是晚期的紅系祖細胞,稱為紅系集落形成單位(colony forming unit-erythroid,CFU-E),它們在體外培養中只能形成較小的集落。晚期紅系祖細胞對BPA不敏感,但主要接受促紅細胞生成素(erythropoietin,EPO)的調節。促紅細胞生成素是一種熱穩定(80℃)的糖蛋白,分子量為34000。當組織中氧分壓降低時,血漿中的促紅細胞生成素的濃度增加,它促進紅系祖細胞向前體細胞分化,又加速這些細胞的增殖,結果使骨髓中能合成血紅蛋白的幼紅細胞數增加,網織紅細胞加速從骨髓釋放。早在本世紀50年代,動物實驗已顯示了促紅細胞生成素活性的存在,以後又確定促紅細胞生成素主要由腎組織產生。切除雙腎後,血漿中促紅細胞生成素的濃度急劇降低。用分子生物學手段進一步證明,從腎組織細胞中已提取出編碼促紅細胞生成素的Mrna 和Cdna,還確定促紅細胞生成素和mRNA和cDNA,還確定促紅細胞生成素基因定位在7號染色體上。近年來有跡象提示人類的某些血液病,如再生障礙性貧血是紅系祖細胞促紅細胞生成素受體有缺陷所致(圖3-2)。
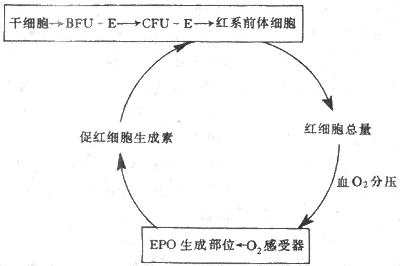
圖3-2 EPOA調節紅細胞生成的反饋調節環
促紅細胞生成素主要由腎組織產生,但腎外,如肝臟,也有小量生成。晚期腎病患者,腎臟產生EPO已基本停止,但體內仍有小量EPO促使骨髓繼續產生紅細胞。
其他一些激素,包括雄激素、甲狀腺激素和生長激素,都可增強促紅細胞生成素的作用;雌激素則有抑制紅細胞生成的作用。這可能是男性的紅細胞數和血紅蛋白量高於女性的原因。
| 關於「生理學/紅細胞生理」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |