營養學/肥胖的病理生理
| 醫學電子書 >> 《臨床營養學》 >> 肥胖 >> 肥胖的病理生理 |
| 臨床營養學 |
|
|
|
由於肥胖的主要發生地點是在機體脂肪組織,所以有關這一節的論述,也就以機體脂肪再重複。
23.4.1 脂肪組織的作用、結構和成分
脂肪組織是結締組織的一種特殊形式,是機體較大的組織之一。在正常成年男性,它佔總體重的15~20%;在正常成年女性,則佔總體重的10~25%。它分布於機體某些特定部位,如皮下、網膜、腸系膜、腹膜後、胸腔縱隔障和胸腹漿膜下等處,起著保護和機械支持的作用,並以TG的形式儲存能量(這是機體最有效、最緊湊的儲能方式),參與糖和脂代謝及體內能量平衡的調節。其中尤以皮下脂肪組織為機體的最大脂肪庫,其儲脂量約佔總體脂量的一半。脂肪組織並不象通常所設想的那樣是一種「沒有多少生活力的組織」,它具有與常代謝活躍組織所具有的相同的全套亞細胞結構。在神經內分泌因素影響下,其TG的合成和分解代謝極為活躍。一些研究表明,當機體進行3H以上的有氧代謝活動,其所需能量幾乎70%要由脂肪組織來提供,而且隨著時間的延長,這一比例還會加大。因此,一定數量的脂肪組織的存在,不僅不是機體的一種「額外負擔」,而恰恰正是機體具有良好身體素質的表現,是機體維持長時間有氧代謝活動的耐力所必需。
脂肪組織主要由脂肪細胞所組成。此外,尚含有少數纖維母細胞和少量細胞間膠原物質。脂肪細胞就處於支持這些細胞及通過這個細胞群的毛細血管和神經纖維的膠原結構之中。脂肪細胞就處於與毛細血管網一道的較原始的原發細胞;原發細胞分化為儲存脂肪的脂肪細胞是受營養和許多內分泌的因素所控制的。
脂肪組織含脂量為60~80%;其所含脂肪都存在於脂肪細胞之內。脂肪細胞的數目,是由遺傳因素和幼年時的飲食因素所控制;而中、晚年時的攝能狀況,則主要影響已有脂肪細胞體積的大小。當前一般認為,正常人全身脂肪細胞數為26.8±1.8×109;但這只是一項參考數值。人體脂肪細胞較為準確的正常數值之所以迄今尚未確定,這是因為:①用目前的方法,一個脂肪細胞一定要在含有最少量的脂質時才能與其它結締組織細胞相區別而能以作出鑒定;②目前所用方法在計算機體脂肪細胞總數時,假定全身各部脂肪細胞的平均直徑是相同的,這又與客觀事實不相符合;③因雌激素水平的不同,正常男性與女性,不僅其體脂含量而且其脂肪細胞數也是不同的,單一數值顯然難以代表。
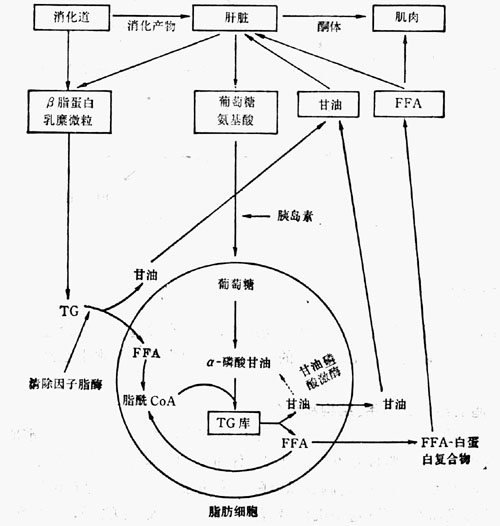
圖23-2 脂肪代謝的某些特徵示意圖
脂肪細胞是機體最大的細胞,正常人皮下脂肪細胞的平均直徑為67~98um,其每一肪細胞的平均含量約0.6ug。含脂質的脂肪細胞在構造上不同於其它細胞的最大特點,主要是由TG組成的一個的中心脂肪滴,將細胞推移到一旁,至少有兩個脂肪區域,一個是由許多快速轉換的細胞質顆粒所組成,另一個則是前者的最終儲存庫,即轉換緩慢的中心性大脂肪滴。脂肪細胞的主要功能是合成、運轉和儲TG。這些功能受神經、激素和酶系所控制,並以FFA形式釋放入血液,以供其它組織的能量需要。脂肪細胞對TG的儲存和釋放,完全取決於機體營養與代謝的需要;脂肪代謝的某些特徵,如上圖所示(圖23-2)。葡萄糖是通過胰島素調節。胰島素不僅促進清除因子脂酶的活性增加,從而增強血漿中脂肪向脂肪細胞內轉移;而且還促進脂肪細胞對葡萄糖的攝取以及對TG的合成。所以,血漿中胰島素的濃度,顯然是決定脂肪組織中脂肪積聚的重要因素。
在機體能量出超的條件下,脂肪組織內所儲存的TG經激素敏感的脂酶的作用而釋放FFA;從脂肪細胞內彌散出來的FFA與作為載體的白蛋白結合或直接進入其他細胞,以供其他組織的能量需要。現在已知,機體大多數組織均可利用FFA,在許多組織中FFA還是一種優先的能源物質。體內絕大多數的激素,如腎上腺素、去甲腎上腺素、生長激素、促腎上腺皮質激素、促甲狀腺激素、黃體生成素、催乳素、胰高血糖素及腸促胰液肽等等,都屬於促進脂肪組織釋放FFA的激素。
23.4.2 脂肪組織中脂肪儲存與釋放的機理
在正常情況下,脂肪組織中的脂肪含量極其恆定,這意味著控制脂肪合成和儲存的反應與控制脂肪分解和動和的反應處於動態平衡之中。為了弄清肥胖的形成,需要複習一下脂肪組織儲存脂肪的調節過程(圖23-3)。
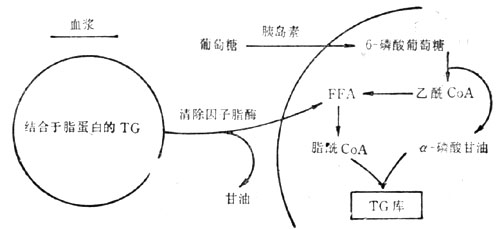
圖23-3 脂肪組織中儲脂途徑示意圖
由上圖可知,脂肪組織儲存脂肪是通過以下兩條途徑:第一,血漿中的脂肪轉移進入脂肪細胞。結合於β-脂蛋白和乳糜微粒的肝源性和腸源性的TG,經秘細血管內皮細胞表面之清除因子脂酶的水解而成為甘油和FFA;接著在酶的調節下FFA進入脂肪細胞,並重新合成TG而儲存起來。為了這個途徑能順利和有效地進行,葡萄糖的攝取和代謝是必需的因為分解的激素,故統稱為「脂解激素」;它們通過刺激脂肪細胞表面的特異感受器,激活與膜結合的腺苷酸環化酶,使細胞內環腺苷酸(cAMP)的濃度增加,進而激活素敏感的脂酶,於是導致FFA的釋放。第二,胰島素則為抑制脂肪分解的激素;它以下四種方式限制FFA的釋放:①抑制腺苷酸環化酶;②激活cAMP磷酸二酯酶,從而促進cAMP的分解;③減慢FFA從脂肪細胞外流;④促進脂肪細胞對葡萄糖的攝取和對FFA的酯化。其所涉及的具體反應如下圖所示(圖23-4)。
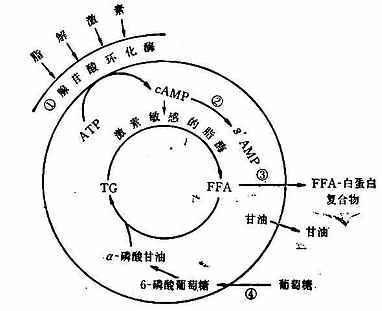
圖23-4 脂肪細胞中脂肪的動員
脂肪細胞的數目,年齡愈小愈多;在20歲以後所發生的肥胖,則幾乎全由於原有脂肪細胞的肥大所致。這說明,除了遺傳因素之外,兒少時期的能量入超,也是一部分人之所以有較多的脂肪細胞的原因。
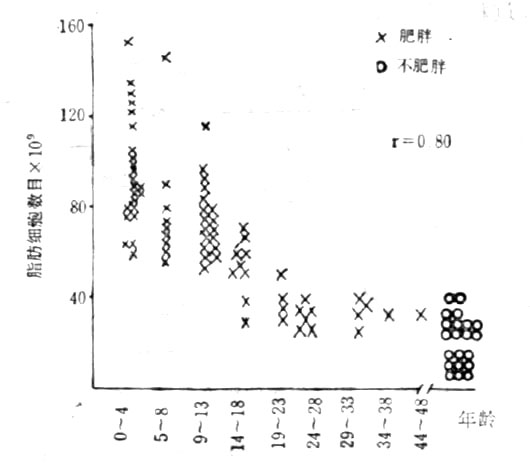
圖23-5 肥胖與不肥胖者的脂肪細胞與年齡的關係
凡因脂肪細胞體積擴大所致的肥胖,通常稱為肥大性肥胖;而因脂肪細胞數目增加所致的肥胖,則稱增生性肥胖。Widdowson等指出,肥胖一旦形成,不論脂肪細胞有多少,其維持低體重的成功率都是低的。尤其是那些缺乏醫學知識的肥胖者,其成功率就更低。臨床經驗還表明,增生性肥胖較肥大性肥胖尤為難治,特別是那些在母體內及剛出生時就已過重的嬰兒。
由上可見,脂肪組織對脂肪的儲存與動用,是在營養、激素和酶系的控制下進行的;而胰島素在調節這兩個過程中所起作用成為重要。
23.4.3 肥胖過程脂肪組織的變化
當肥胖發生和發展時,隨著體脂積聚,脂肪細胞含脂量漸增,其體積也明顯擴大。以皮下脂肪為例,每一細胞的平均含脂量可由原來的0.6ug增至0.91~1.36ug,其平均直徑則可由原來的67~98um增至127~134um。尤其當肥胖的發生與發展過程是緩慢而持續時,有僅可發生更為肥大的脂肪細胞,而且通過脂肪組織的毛細血管內皮細胞表面還可含更多高活性的清除因子脂酶。同時,某些肥胖者還會具有數目較多的脂肪細胞,其全身脂肪細胞數可達77.0±13.5×109。無論是脂肪細胞體積的擴大或脂肪細胞數目的增多,或者是兩者兼而有之,其結果都是機體脂肪組織的擴增。
Salans等對78名肥胖者和21名非肥胖者作了對比觀察,結果(圖23-5)表明:發生肥胖嬰兒期明顯肥胖、青春發育期肥胖,有這樣長期持續肥胖史的成年肥胖者,往往是脂肪細胞的肥大和增生同時並存,其治療難度也就更大不僅不易取得療效,而且還能於鞏固療效。據Lioyd報導,有這樣長期持續肥胖史的肥胖者,在整個成年肥胖者中佔1/3;而其餘的2/3,則分別發生於一生中各個有生理性增加體脂的階段。而Abrham等對10~13歲肥胖和正常兒童各100名(男女對半)作了長期的對比觀察,歷時20年,結果肥胖兒男性43/50、女性40/50變成肥胖成人,正常兒則男性21/50、女性9/50變成肥胖成人。由上可見,從胎兒時期開始,直至一生中各個有生理性增加體脂的階段,及早地採取措施預防肥胖,也就至關緊要。
23.4.4 TBAT之狀況與機體胖瘦的關係
在人和多數哺乳動物(特別是嬰兒和新生仔畜)的頸、肩、腋窩和背部肩胛間,還存在一種特殊類型的脂肪組織。它的結構特點是在脂肪細胞中除含大量分散的TG滴之外,還含有大量粒線體,血液供應豐富,由於其粒線體中細胞色素多,而使之呈現棕色,故通常也就稱之為「棕色脂肪組織(IBAT)」。與其相對應,前述之普通脂肪組織則就稱之為「白色脂肪組織」。
IBAT的主要功能是產熱。成年機體遭受刺激,通常可通過戰慄和化學兩種途徑產熱;可是,嬰兒及新生仔畜由於還不會通過戰慄途徑產熱,其所需之熱幾乎全要靠化學途徑來供應。除Na+-K+泵外,IBAT就是機體化學產熱的又一「重要裝置」;在某些小動物甚至其所需總熱量的一半是由IBAT所提供。故一定數量質量的IBAT的存在,對機體適應寒冷刺激具有重要意義。IBAT的產熱作用,是在交感神經與一種分子量為32000的蛋白質類「脫偶合因子」的控制下進行的。交感神經有豐富的神經末梢支配這種組織;而這種組織的呣粒體中則有大量的為其所特有的「脫偶合因子」的存在。寒冷刺激引起交感神經興奮,其末梢就釋放兒茶酚胺,於是就通過cAMP而使激素敏感的脂酶活化,促使TG水解為FFA。這時,又由於該組織中有大量細胞色素及其所特有的「脫偶合因子」的存在,同時高水平的FFA亦有脫偶合作用,致使FFA被氧化,但偶合則很少,從而能量也就很少產生ATP,而大部分以熱能的形式放出。再加上,脫偶合結果導致了ATP~ADP比值的減少,而較多的ADP又使氧化磷酸化及呼吸作用加快,這樣也就更使熱生成作用增強。
晚近通過對IBAT產熱機理的深入研究,發現IBAT之數與質的狀況與機體的胖瘦之間有著密切的相關。就目前所知,消瘦、正常、肥胖三者相比,除了機體所含白色脂肪組織明顯順序遞增而IBAT明顯順序遞減外,其IBAT粒線體中所含的特有的「脫偶合因子」的數量和活性也明顯順序遞減。正常者由於其白色脂肪組織與IBAT之間比例為平衡,而且「脫偶合因子」的數量和活性也較適中,能對產熱和產生ATP進行有效的調節,故在通常情況下能使機體能量得以維持正常的平衡,即能使之不過多以熱生成的形式而被消耗,又能使之不過多轉化為脂肪而在體內積聚。至於消瘦者,則往往是因為其IBAT含量較多,以及其「脫偶膈因子」的數量和活性較高,致使其能量較多地以熱生成的形式被消耗了,所以在一般情況下也就難以有較多的體脂積聚;反之,肥胖者則由於其所含IBAT量少或功能障礙,致使產熱這一有效的調節方式失靈,所以引起能量的入超,其大部分就轉化為脂肪而積聚起來。
| 關於「營養學/肥胖的病理生理」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |