醫學微生物學/機體抗病毒免疫
| 醫學電子書 >> 《醫學微生物學》 >> 病毒的感染與免疫 >> 機體抗病毒免疫 |
| 醫學微生物學 |
|
|
|
機體抗病毒感染免疫應答包括非特異性免疫與特異性免疫。前者指獲得性免疫力產生之前,機體對病毒初次感染的天然抵抗力,主要為單核吞噬細胞、自然殺傷細胞及干擾素等的作用。後者指抗體介導的和細胞介導的抗病毒作用。本節中干擾素將單獨闡述。
目錄 |
一、非特異性因素的抗病毒作用
(一)機械和化學屏障
皮膚為鱗狀上皮組織,是陰止病毒感染的良好屏障;呼吸道粘膜細胞纖毛的反向運動是一種保護機制,當流感、副流感病毒感染破壞了粘膜細胞時,易發生繼發感染;胃酸對病毒有滅活作用,有囊膜病毒一般不能通過消化道感染,多數無囊膜腸道病毒是耐酸的;血腦屏障和胎盤屏障可阻止大多數病毒感染腦細胞和胎兒。
(二)單核吞噬細胞和自然殺傷細胞
單核吞噬細胞,尤其是固定或遊走的巨噬細胞吞噬並消化大分子異物,抗體或補體的活性成分起調理吞噬作用;IFN-r活化的巨噬細胞增強殺滅病毒的能力。
自然殺傷細胞(NK細胞)在無抗原刺激的情況下,通過非抗體依賴的方式自然殺傷腫瘤細胞及病毒感染的細胞,是機體抗腫瘤、抗病毒的重要防線,在血中占淋巴細胞的10%。此外,NK細胞尚具有抗體依賴性細胞介導的細胞毒作用(ADCC)。
病毒感染部位引起炎細胞浸潤,中性粒細胞,而後是巨噬細胞、淋巴細胞聚集,導致局部氧利用和酸性產物增加,不利於病毒複製,限制病毒的擴散。巨噬細胞產生IL-1及IFN引起機體發熱反應,當超過37℃時大多數病毒複製受到壓抑,因此發熱是一種保護性機制,使用藥物強制退熱將延長病毒感染的康復時間。然而過強的炎症反應和高燒也是病情加重的因素。
(四)年齡與生理狀態
嬰兒有來自母體的lgG及母乳中的lgA,6個月內較少病毒性感染;兒童脊髓灰質炎病毒通常引起輕型或亞臨床型感染,然而在成年人則往往是麻痹型感染;老年人因免疫力下降,患帶狀皰疹較為嚴重。妊娠期患B型肝炎有時病情惡化,可能與內分泌變化有關;使用激素治療雖可控制高燒及過強的炎症反應,但往往加重單純皰疹和水痘帶毒等感染的病情。
動物實驗證明遺傳因素影響病毒感染的抵抗力或敏感性。近交系小鼠接種少量單純皰疹病毒便可致命,而其他小鼠可耐受較大劑量病毒的攻擊不出現症状,涉及的基因與MHC有關。在人體中難以做出評價,但隨著免疫應答基因的深入研究有助於闡明這一問題。
許多病毒有限定的宿主範圍,種屬免疫可能取決於宿主細胞是否具有相應的受體,如脊液灰質炎病毒的受體僅存在於人和靈長類動物中,因而其他動物具有天然的種屬免疫力;人類免疫缺陷病毒、肝炎病毒也是如此。相反,狂犬病毒可感染多種溫血動物。
二、抗體介導的抗病毒作用
(一)中和抗體理與非中和抗體
具有吸附穿入作用的病毒表面抗原所誘生的抗體,稱之為中和抗體。活病毒與中和抗體結合,導致病毒喪失感染力,稱為中和反應。結合這種抗體的病毒不能再吸附和穿入易感宿主的細胞。如抗流感病毒血凝素抗原的抗體,為中和抗體,具有免疫保護作用。血流中特異性lgM出現於病毒感染的早期,lgG出現較晚,它們都能抑制病毒的局部擴散和清除病毒血症,並能抑制原發病灶中病毒播散至其他易感組織和器官(靶器官)。粘膜表面分泌型lgA的出現比血流中lgM稍晚,它是呼吸道和腸道抵抗病毒的重要因素。而補體能明顯地加強中和抗體的作用。
不具有吸附穿入作用的病毒表面抗原及病毒顆粒內部抗原所誘生的抗體,稱之為非中和抗體。如抗流感病毒神經氨酸酶的抗體,不能阻止病毒吸附穿放敏感細胞,但可與病毒表面神經氨酸酶結合,易被吞噬清除;抗流感病毒核蛋白的抗體,沒有免疫保護作用。
(二)抗體介導的抗病毒作用方式
1.中和抗體和病毒表面抗原結合,導致病毒表面蛋白質構型的改變,阻止其吸附於敏感細胞。中和作用是機體滅活游離病毒的主要方式。
2.病毒表面抗原(如流感病毒的血凝素和神經氨酸酶)與相應的抗體(中和抗體或非中和抗體)結合時,易被吞噬清除。
3.病毒表面抗原和相應的抗體結合後,激活補體,導致有囊膜的病毒裂解。
4.感染細胞表面表達的病毒抗原與相應抗體結合後,或通過ADCC作用或通過激活補體,使靶細胞溶解。
三、細胞介導的抗病毒作用
(一)細胞毒T淋巴細胞(CTL)的抗毒作用
雖然NK細胞和活化的巨噬細胞有殺傷靶細胞的作用,但破壞病毒感染的靶細胞主要靠CTL,CTL由CD+8的和CD+4的殺傷細胞組成.CD+8的淋巴細胞約佔成熟T淋巴細胞的30%,包括大多數CTL和抑制性T細胞(Ts).當病毒抗原與宿主細胞MHC-1類抗原一起提呈給CD+8的CTL時才能增殖為活化的殺傷細胞,它們殺傷靶細胞受MHC-1類抗原的限制。CTL殺傷靶細胞的機制可能是釋放胰蛋白酶樣的絲氨酸蛋白酶和細胞毒性淋巴因子(如穿孔素-Perfarin)等,導致靶細胞溶解。CD+4的淋巴細胞約佔成熟T淋巴細胞的70%,主要為輔助性T細胞(Th),尚有少數CTL。它們的活化與殺傷功能受細胞表面相應的MHC-Ⅱ類抗原的限制。
Th和Ts是重要的免疫調節細胞,在血中正常比率為2:1。病毒抗原大多為胸腺依賴性抗原,Th和Ts分別從正向和反向調節機體抗病毒免疫應答的強弱程度,影響抗體介導的和細胞介導的免疫保護或免疫損傷作用。
(二)細胞因子的抗病毒作用
細胞毒效應細胞(CTL、NK及活化的巨噬細胞)與靶細胞結合後,通過釋放出的細胞毒性物質殺傷病毒感染的靶細胞,前已述及。近年來在小鼠體內實驗中證實,活化的T細胞釋放的LL-2促使T細胞、NK細胞、單核巨噬細胞在病毒感染的部位大量聚集,釋放IFN-γ、IFN-α並形成足夠的濃度,從而在基因轉錄、翻譯兩個水平上抑制病毒大分子的合成,並誘導鄰近正常細胞建立抗病毒狀態。因此,它們在機體抗病毒免疫中是極為重要的效應分子。
四、干擾素的抗病毒作用
當二種病毒感染同一細胞時,一種病毒可以增殖,而另一種病毒則被抑制,這種現象稱為干擾現象(Interference),這種能干擾病毒增殖的物質稱為干擾素(Interferon,IFN)。IFN是在誘生劑和某些細胞因子的作用下,由細胞基因編碼產生的一組蛋白質,具有高度活性和多種功能。IFN無病毒特異性,一種病毒誘生的IFN對其病毒也有效,但有種屬特異性,小鼠產生的的IFN在人體內無效。現知人的干擾素有3種:IFN-α即白細胞干擾素,IFN-β即成纖維細胞干擾素,IFN-γ即免疫干素(T淋巴細胞產生的一種細胞因子);前二種屬Ⅰ型,後一種屬Ⅱ型。編碼Ⅰ型IFN的基因位於人類第9對染色體上,編碼Ⅱ型IFN的基因位於第12對染色體上。
(一)干擾素的誘生
在正常情況下,基因處於靜止狀態,干擾素的產生受到抑制。如有病毒感染或非病毒性誘生劑(如人工合成的雙鏈聚肌胞,Poly I:C)作用於細胞膜上,激活干擾素編碼基因,即開始轉錄干擾素的mRNA,再轉譯為干擾素蛋白。因此誘生的干擾素很快釋放到細胞外,作用於鄰近的未受感染的細胞膜受體上,使細胞建立抗病毒狀態。
(二)干擾素作用的機理
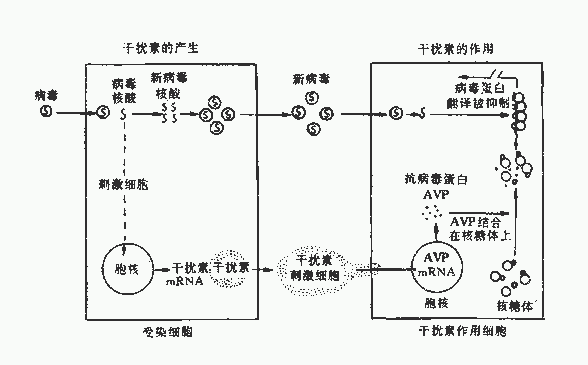
圖22-4 干擾素的產生及其作用機制
干擾素首先作用於鄰近未受感染的細胞膜上的干擾素受體系統,該系統由神經節苷脂組成的結合位點和一個可能由糖蛋白組成的激活位點所組成。現已知Ⅰ型干擾素受體基因在人染色體G21長臂上,Ⅱ型干擾素受體基因位於第6對染色體上,這就決定了IFN具有一定的程屬特異性。當IFN與受體結合後,產生一種特殊的因子,使抗病毒蛋白(AVP)基因解除抑制,轉錄並翻譯出AVP,主要是蛋白激酶、2ˊ-5ˊA合成酶、磷酸二酯酶,這些酶與發揮抗病毒活性有密切關係。其中二種酶須經雙股RNA及ATP激活,一為蛋白激酶,激活後可使合成蛋白質的起動因子2(eIF-2)同磷酸化而失活,抑制蛋白質合成;另一種為2ˊ-5ˊA合成酶,活化後催化合成2ˊ-5ˊ寡腺苷酸,再激活潛在的核酸內切酶,使病毒mRNA降解,抑制病毒蛋白合成。此外,磷酸二酯酶能降解2ˊ-5ˊA,又能除去tRNA的CCA末端,抑制蛋白合成。
干擾素在病毒學、免疫學、細胞學及分子遺傳學等領域,均有理論與實際意義。干擾素製劑已用於治療一些病毒感染(如慢性B型肝炎、單純皰疹病毒毒性角膜炎、帶狀皰疹及呼吸道病毒感染)和惡性腫瘤(如成骨肉瘤等),似都有一定療效。在病毒感染時,干擾素的產生較特異性抗體早,因此,對於阻止病毒病的發生及病毒病患者的康復可能起重要作用。
表22-9 各種免疫因素及其抗病毒作用
| 抗病毒作用 | 免疫因素 |
| 滅活和清除游離病毒 | 1.主要靠中和抗體 2.補體能增加抗體的中和活性 3.巨噬細胞清除病毒 |
| 保護鄰近細胞免於感染、抑制病毒的增殖 | 干擾素 |
| 破壞感染病毒的靶細胞: 病毒增殖中斷 |
1.抗體加補體的溶細胞作用 2.CTL及某些細胞因子 3.抗體依賴性細胞介導的細胞毒作用(ADCC)(NK細胞) 4.激活的巨噬細胞 5.自然殺傷性(NK)細胞 |
| 關於「醫學微生物學/機體抗病毒免疫」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |