生物化學與分子生物學/參與蛋白質生物合成的物質
| 醫學電子書 >> 《生物化學與分子生物學》 >> 蛋白質的生物合成 >> 參與蛋白質生物合成的物質 |
| 生物化學與分子生物學 |
|
|
|
目錄 |
(一)合成原料
自然界由mRNA編碼的胺基酸共有20種,只有這些胺基酸能夠作為蛋白質生物合成的直接原料。某些蛋白質分子還含有羥脯氨酸、羥賴氨酸、γ-羧基谷氨酸等,這些特殊胺基酸是在肽鏈合成後的加工修飾過程中形成的。
(二)mRNA是合成蛋白質的直接模板
原核細胞中每種mRNA分子常帶有多個功能相關蛋白質的編碼信息,以一種多順反子的形式排列,在翻譯過程中可同時合成幾種蛋白質,而真核細胞中,每種mRNA一般只帶有一種蛋白質編碼信息,是單順反子的形式。mRNA以它分子中的核苷酸排列順序攜帶從DNA傳遞來的遺傳信息,作為蛋白質生物合成的直接模板,決定蛋白質分子中的胺基酸排列順序。不同的蛋白質有各自不同的mRNA,mRNA除含有編碼區外,兩端還有非編碼區。非編碼區對於mRNA的模板活性是必需的,特別是5』端非編碼區在蛋白質合成中被認為是與核糖體結合的部位。見圖18-2。
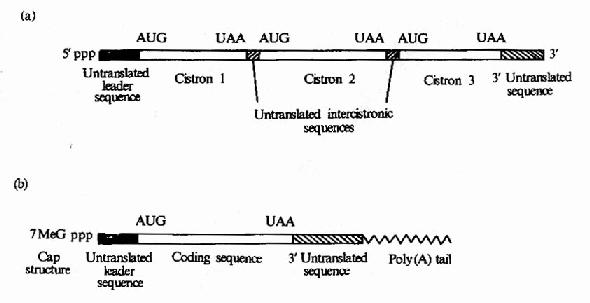
圖18-2 (a)原核生物mRNA)為多順反子(b)真核生物mRNA為單順反子
mRNA分子上以5'→3'方向,從AUG開始每三個連續的核苷酸組成一個密碼子,mRNA中的四種鹼基可以組成64種密碼子。這些密碼不僅代表了20種胺基酸,還決定了翻譯過程的起始與終止位置。每種胺基酸至少有一種密碼子,最多的有6種密碼子。從對遺傳密碼性質的推論到決定各個密碼子的含義,進而全部闡明遺傳密碼,是科學上最傑出的成就之一,科學家們設計了十分出色的遺傳學和生物化學實驗,於1966年編排出了遺傳密碼字典。見表18-1。
表18-1 胺基酸的密碼(code)
| 5』末端(第1位鹼基) | 中間鹼基(第二位鹼基) | 3』末端(第三位鹼基) | |||
| U | C | A | G | ||
| U | 苯丙(Pne)F | 絲(Ser)S | 酪(Tyr)Y | 半胱(Cys)C | U |
| 苯內(Pne) | 絲(Ser) | 酪(Tyr) | 半胱(Cys) | C | |
| 亮(Leu)L | 絲(Ser) | 終止信號 | 終止信號 | A | |
| 亮(Leu) | 絲(Ser) | 終止信號 | 色(Trp) | G | |
| C | 亮(Leu) | 脯(Pro)P | 組(His)H | 精(Arg)R | U |
| 亮(Leu) | 脯(Pro) | 組(His) | 精(Arg) | C | |
| 亮(Leu) | 脯(Pro) | 谷胺(Gin)Q | 精(Arg) | A | |
| 亮(Leu) | 脯(Pro) | 谷胺(Gin) | 精(Arg) | G | |
| A | |||||
| 異亮(ILe)I | 蘇(Thr)T | 天胺(Asn)N | 絲(Ser)S | U | |
| 異亮(ILe) | 蘇(Thr) | 天胺(Asn) | 絲(Ser) | C | |
| 異亮(ILe) | 蘇(Thr) | 賴(Lys)K | 精(Arg)R | A | |
| *蛋(Met)M(起動信號) | 蘇(Thr) | 賴(Lys) | 精(Arg) | G | |
| G | |||||
| 纈(Val)V | 丙(Ala)A | 天(Asp)D | 甘(Gly)G | U | |
| 纈(Val) | 丙(Ala) | 天(Asp) | 甘(Gly) | C | |
| 纈(Val) | 丙(Ala) | 谷(Glu)E | 甘(Gly) | A | |
| 纈(Val) | 丙(Ala) | 谷(Glu) | 甘(Gly) | G | |
遺傳密碼具有以下幾種特點:
(1)起始碼與終止碼(Initiation codon and termination codon):
密碼子AUG是起始密碼,代表合成肽鏈的第一個胺基酸的位置,它們位於mRNA5′末端,同時它也是蛋氨酸的密碼子,因此原核生物和真核生物多肽鏈合成的第一個胺基酸都是蛋氨酸,當然少數細菌中也用GUG做為起始碼。在真核生物CUG偶爾也用作起始蛋氨酸的密碼。密碼子UAA,UAG,UGA是肽鏈成的終止密碼,不代表任何胺基酸,它們單獨或共同存在於mRNA3』末端。因此翻譯是沿著mRNA分子5′→3′方向進行的。
(2)密碼無標點符號:兩個密碼子之間沒有任何核苷酸隔天,因此從起始碼AUG開始,三個鹼基代有一個胺基酸,這就構成了一個連續不斷的讀框,直至終止碼。如果在讀框中間插入或缺失一個鹼基就會造成移碼突變,引起突變位點下游氨基排列的錯誤。
(3)密碼的簡併性(Degemeracy):
一種胺基酸有幾組密碼子,或者幾組密碼子代表一種胺基酸的現象稱為密碼子的簡併性,這種簡併性主要是由於密碼子的第三個鹼基發生擺動現象形成的,也就是說密碼子的專一性主要由前兩個鹼基決定,即使第三個鹼基發生突變也能翻譯出正確的胺基酸,這對於保證物種的穩定性有一定意義。如:GCU,GCC,GCA,GCG都代表丙氨酸。
(4)密碼的通用性:
大量的事實證明生命世界從低等到高等,都使用一套密碼,也就是說遺傳密碼在很長的進化時期中保持不變。因此這張密碼錶是生物界通用的。然而,出乎人們預料的是,真核生物粒線體的密碼子有許多不同於通用密碼,例如人粒線體中,UGA不是終止碼,而是色氨酸的密碼子,AGA,AGG不是精氨酸的密碼子,而是終止密碼子,加上通用密碼中的UAA和UAG,粒線體中共有四組終止碼。內部甲硫氨酸密碼子有兩個,即AUG和AUA;而起始甲硫氨酸密碼子有四組,即AUN。
密碼子結構與胺基酸側鏈析性之間也有一定關係。①胺基酸側鏈極性性質在多數情況下由斷面子的第二個鹼基決定。第二個鹼基為嘧啶(Y)時,胺基酸側鏈為非極性,第二個鹼基為嘌呤時,胺基酸側鏈則有極性。②當第一個鹼基為U或A,第二個鹼基為C,第三個鹼基無特異性時,所決定的胺基酸側鏈為極性不帶電。③當第一個鹼基不是U,第二個鹼基是P時,胺基酸側鏈則帶電。在此前提下,若是一個是C或A時,表示帶正電的胺基酸,第一、二個鹼基分別是G、A時,此種氨革酸帶負電,但上述關係也有個別例外。
一種胺基酸由多種密碼子所編碼的事實使人想到:同一種胺基酸的一組密碼子的使用頻率是否相同?許多實驗證實,在原核生物和高等真核生物中同一組密碼子的使用頻率是不相同的。高頻密碼子多出現在那些表達量高的蛋白質基因中,例如,核糖體蛋白質基因,RecA蛋白質基因等。表18-2。這種使用頻率與細胞內一組tRNA中的不同tRNA含量有關。
(三)tRNA是胺基酸的運載工具:
tRNA在蛋白質生物合成過程中起關鍵作用。mRNA推帶的遺傳信息被翻譯成蛋白質一級結構,但是mRNA分子與胺基酸分子之間並無直接的對應關係。這就需要經過第三者「介紹」,而tRNA分子就充當這個角色。tRNA是類小分子RNA,長度為73-94個核苷酸,tRNA分子中富含稀有鹼基和修飾鹼基,tRNA分子3』端均為CCA序列,胺基酸分子通過共價鍵與A結合,此處的結構也叫胺基酸臂。每種胺基酸都有2-6種各自特異的tRNA,它們之間的特異性是靠氨基醯tRNA合成酶來識別的。這樣,攜帶相同胺基酸而反密碼子不同的一組tRNA稱為同功tRNA,它們在細胞內合成量上有多和少的差別,分別稱為主要tRNA和次要tRNA。主要tRNA中反密碼子識別tRNA中的高頻密碼子,而次要tRNA中反密碼子識別mRNA中的低頻密碼子。每種胺基酸都只有一種氨基醯tRNA合成酶。因此細胞內有20種氨基醯tRNA合成酶。
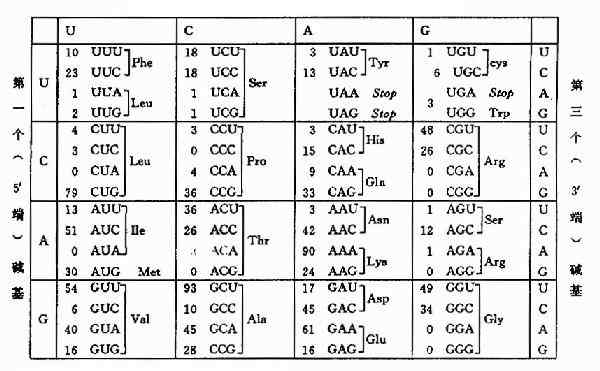
圖18-3 密碼子和反密碼子的相互作用
tRNA分子中還有一個反密碼環,此環上的三個反密碼子的作用是與mRNA分子中的密碼子靠鹼基配對原則而形成氫鍵,達到相互識別的目的。但在密碼子與反密碼子結合時具有一定擺動性,即密碼子的第3位鹼基與反密碼子的第1位鹼基酸對時並不嚴格,見圖18-3。配對擺動性完全是由tRNA反密碼子的空間結構所決定的。反密碼的第1位鹼基常出現次黃嘌呤I,與A、C、U之間皆可形成氫鍵而結合,這是最常見的擺動現象。這種擺動現象使得一個tRNA所攜帶的胺基酸可排列在2-3個不同的密碼子上,因此當密碼子的第3位鹼基發生一定程度的突變時,並不影響tRNA帶入正確的胺基酸。
表18-3 反密碼與密碼鹼基配對時的搖擺現象
| 反密碼第1位鹼基 | A | C | G | U | 1 |
| 密碼第3位鹼基 | U | G | C,U | A,G | A,C,U |
在蛋白質生物合成過程中,特異識別mRNA上起始密碼子的tRNA被稱為起始tRNA,它們參加多肽鏈合成的起始,其它在多肽鏈延伸中運載胺基酸的tRNA,統稱為延伸tRNA。
(四)核糖核蛋白體
核蛋白體是由rRNA和幾十種蛋白質組成的亞細胞顆粒,位於胞漿內,可分為兩類,一類附著於粗面內質網,主要參與白蛋白、胰島素等分泌性蛋白質的合成,另一類游離於胞漿,主要參與細胞固有蛋白質的合成。核糖體是細胞中的主要成分之一,在一個生長旺盛的細菌中大約不20000個核糖體,其中蛋白質占細胞總蛋白質的10%,RNA占細胞總RNA的80%。
任何生物的核糖體都是由大、小兩個亞基組成,現將大腸桿菌核糖體和大鼠肝細胞核糖體的蛋白質組分和RNA組成列表於18-4。1968年已在體外對大腸桿菌小亞基進行了自我裝配研究,加入16s rRNA和21種蛋白質,即可形成有天然活性的30s小亞基。通過這些研究使人們能夠進一步認識小亞基和大亞基中rRNA與蛋白質的特異功能。核糖體是高度複雜的體系,它的任何個別組分或局部組分都不能起整體的作用,因此必須研究核糖體中蛋白質和RNA的空間結構和位置,才能更完全地了解蛋白質合成的具體過程。過去一直認為rRNA主要起著結構上的作用,蛋白質發揮催化功能,但現在認為rRNA與蛋白質共同的構成的核糖體功能區是核蛋白體表現功能的重要部位,如GTP酶功能區,轉肽酶功能區以及mRNA功能區等等。
表18-4 核蛋白體的組成及特性
| 來源 | 直徑(毫微米) | 重量(道爾頓) | 含rRNA(%) | 含蛋白質(%) | 沉降係數 | 亞基 | 含rRNA | 含蛋白質種數 | 每個細胞內含有的個數 | |
| 種類 | 分子量 | |||||||||
| 真核細胞胞液 | 20~22 | 3.6×106 | 55 | 45 | 77S~80s | 40S(小) | 18S | -70萬 | ~34 | 106~107 |
| 60S(大) | 5s 5.8S 28S~29S |
3萬 4萬 140~180萬 |
~40 | |||||||
| 原核細胞胞液 | 18 | 2.6×106 | 60~65 | 30~35 | 70S | 30S(小) | 16S | 55萬 | ~34 | 1.5×104 |
| 50S(大) | 5s 23S |
4萬 110萬 |
||||||||
註:真核細胞粒線體的核蛋白體組成及特性與原核細胞胞液的相同
核蛋白體作為蛋白質的合成場所具有以下幾種作用:
(1)mRNA結合位點:位於30s小亞基頭部,此處有幾種蛋白質構成一個上的結構域,負丙與mRNA的結合,特別是16srRNA3』端與mRNa AUG之前的一段序列互補是這種結合必不可少的,見表18-5。
表18-5 大腸桿菌mRNA起始密碼上游區域SD序列和16srRNA的互補
| MS2外殼蛋白 MS2複製酶 MS2A蛋白 λCro galE β-內醯氨酶 脂蛋白 核糖體蛋白S12 RNA聚合酶β trpE |
3』HOAU UCCUCCACUAG……5』 5』……UCAACC GGAGUUUGAAUCAUG…3』 5』……CAAACAU GAGGAUUACCCAUG …3』 5』…… UCCU AGGAGGUUUGACCUGUG…3』 5』…… AUGUAC UAAGGAGGUUGUAUG…3』 5』…… AGCCUAAU GGAGCGAAUUAUG…3』 5』…… UAUUGAAA AAGGAAGAGUAUG…3』 5』…… AUCUA GAGGGUAUUAAUAAUG…3』 5』…… AAAACCAGGAGCUAUUUAAUG…3』 5』…… AGCGAGCU GAGGAACCCUAUG…3』 5』…… CAAAAUUAGAGAAUAACAAUG…3』 |
(2)P位點:(peptidyl tRNA site)
又叫做肽醯基tRNA位或給位。它大部分位於小亞基,小部分位於大亞基,它是結合起始tRNA 並向A位給出胺基酸的位置(圖18-4)。
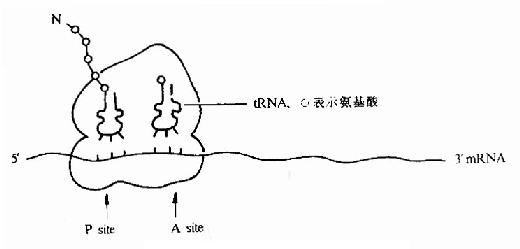
圖18-4 翻譯過程中的核糖體圖解
(3)A位點:(Aminoacyl-tRNA site)
叫做氨基醯 tRNa 位或受位。它大部分位於大亞基而小部分位於小亞基,它是結合一個進入的氨基醯tRNA 的位置(見後節敘述)。
(4)轉肽酶活性部位:
位於P位和A位的連接處。
(5)結合參與蛋白質合成的起始因了(Initiation Factor,IF)、延長因子(ElengationFactor,EF)和終止因子或釋放因子(Release Factor,RF)。
| 關於「生物化學與分子生物學/參與蛋白質生物合成的物質」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |