生理學/由膜的特異受體蛋白質、G-蛋白和膜的效應器酶組成的跨膜信號傳遞系統
| 醫學電子書 >> 《生理學》 >> 細胞的基本功能 >> 細胞的跨膜信號傳遞功能 >> 由膜的特異受體蛋白質、G-蛋白和膜的效應器酶組成的跨膜信號傳遞系統 |
| 生理學 |
|
|
|
這是另一類型的跨膜信號傳遞。最初是從對激素作用機制的研究開始的。60年代在研究腎上腺素引起肝細胞中糖原分解為葡萄糖的作用機制時,發現如果使腎上腺素單獨和分離出的細胞膜碎片相互作用,可以生成一種分子量小、能耐熱的物質,當把這種物質同肝細胞的胞漿單獨作用時,也能引起其中糖原的分解,同腎上腺素作用於完整的肝細胞時有類似的效應。實驗提示,在腎上腺素正常起作用時,它只是作用於肝細胞的膜表面。通過某種發生在膜結構中的過程,先在胞漿中生成一種小分子物質,後者再實現腎上腺素分解糖原的作用。這種小分子物質不久被證明是環-磷酸腺苷(即cAMP,環磷腺苷)。以後又陸續發現,很多其他激素類物質作用於相應的靶細胞時,都是先同膜表面的特異受體相結合,再引起膜內側胞漿中cAMP含量的增加(有時是它的減少),實現激素對細胞內功能的影響。這樣就把cAMP稱作第二信使,這是相對於把激素分子這類外來化學信號看作第一信使而言的。
導致cAMP產生的膜結構內部的過程頗為複雜:它至少與膜中三類特殊的蛋白質有關。第一類是能與到達膜表面的外來化學信號作特異性結合的受體蛋白質,這是一些真正可以稱作受體的物質。目前已用分子生物學的方法證明,它們是一些獨立的蛋白質分子;已經確定的近100種這類受體,都具有類似的分子結構,也屬於同一蛋白質家族:即它們都由約300~400個氨基酸殘基組成,有一個較長的細胞外N-末端,接著在肽鏈中出現7個由22~28個主要為疏水性氨基酸組成的α-螺旋,說明這肽鏈至少要反覆貫穿膜7次,形成一個球形蛋白質分子,還有一段位於膜內側的肽鏈C-末端。目前認為,受體分子中第7個跨膜螺旋是能夠識別、即能結合某種特定外來化學信號的部位;在受體因結合了特異化學信號而激活時,將進而作用於膜中另一類蛋白質,即G-蛋白質。
G-蛋白是鳥苷酸結合蛋白(guaninenucleotide-binding protein)的簡稱,也是存在於膜結構中的一類蛋白質家族,根據它們分子結構中少數氨基酸殘基序列上的不同,已被區分出有數十種,但結構和功能極為相似。G-蛋白通常由α-、β-、和γ-3個亞單位組成;α-亞單位通常起催化亞單位的作用,當G-蛋白未被激活時,它結合了一分子的GDP(二磷酸鳥苷);當G-蛋白與激活了的受體蛋白在膜中相遇時,α-亞單位與GDP分離而又與一分子的GTP(三磷酸鳥苷)結合,這時α-亞單位同其他兩個亞單位分離,並對膜結構中(位置靠近膜的內側面)的第三類稱為膜的效應器酶的蛋白質起作用,後者的激活(或被抑制)可以引致胞漿中第二信使物質的生成增加(或減少)。上述腎上腺素的作用,就是先由激素激活膜上相應的受體後,通過一種稱為Gs(興奮性G-蛋白)的G-蛋白的中介,激活了作為效應器酶的腺苷酸環化酶(圖2-9箭頭1),使胞漿中的ATP生成了起第二信使作用的cAMP(圖2-9中箭頭2)。由於第二信使物質的生成經過多級催化作用,少數幾個膜外化學信號分子同受體的結合,就可能在胞漿中生成數目眾多的第二信使分子,這是這種類型的跨膜信號傳遞的重要特點之一。
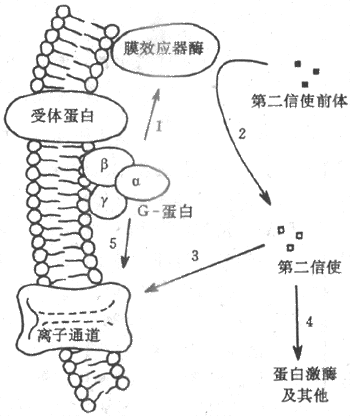
圖2-9 由膜受體-G-蛋白-膜效應器酶組成的跨膜信號
傳遞系統和第二信使類物質的生成
目前發現膜的效應器酶並不只腺苷酸環化酶一種,因而第二信使物質也不只cAMP一種,如近年來還發現,有相當數量的外界刺激信號作用於受體後,可以通過一種稱為Go的G-蛋白,再激活一種稱為磷脂酶C的膜效應器酶,以膜結構中稱為磷脂醯肌醇的磷脂分子為間接底物,生成兩種分別稱為三磷酸醯肌醇(IP3)和二醯甘油(DG)的第二信使,影響細胞內過程,完成跨膜信號傳遞。雖然如此,對應於細胞所能接受的多種刺激和與它們相對應的受體數目而言,膜內G-蛋白、效應器酶和最後生成的第二信使類物質的種類,還是相對地少得多。這說明,上述由膜中蛋白質酶促反應生成第二信使的途徑,具有相當程度的「通用」性質。
由於上述這種跨膜信號傳遞的形式是在研究激素的作用機制時發現的,而且後來發現絕大多數肽類激素都是通過這一形式起作用的,因此曾一度錯誤地認為,這只是激素性化學信號跨膜信號傳遞方式。但近年的資料說明,事實並非如此:在神經遞質類物質中,除了上述氨基酸類遞質外,其餘不論是小分子的經典遞質還是後來發現的數量眾多的神經肽類物質(目前已近50種),都主要是以在突觸後細胞中產生第二信使類物質來完成跨膜信號傳遞的,這些第二信使物質通過在胞漿中的擴散,在膜的內側面作用於某些特殊的離子通道(圖2-9中箭頭3),引起突觸後膜較廣泛而緩慢的電變化。最近證明,在視網膜信號轉換過程中,光量子被作為受體的視色素如視紫紅質(也具有7個跨膜α-螺旋的結構特點)吸收後,也是先激活稱為Gt(轉換蛋白)的G-蛋白,再激活作為效應器的磷酸二酯酶,使視桿細胞外段中cGMP的分解加強,最後使光刺激轉變為外段膜的電變化(見第九章)。
上述兩種主要的跨膜信號傳遞方式的作用過程,有以下幾點值得注意。第一,這兩種作用形式並不是絕對分離的,兩者之間可以互相影響或在作用上有交叉。一些第二信使類物質可以調節某些電壓門控通道和化學門控通道蛋白質的功能狀態;而且被某種受體激活了的G-蛋白,有的不通過第二信使就能直接作用於膜結構中的通道結構(圖2-9中的箭頭5),如上述Gs激活時可以直接打開Ca2+通道。第二,對於許多外來化學信號分子,並不是一種化學信號只能作用於兩種跨膜信號傳遞系統中的一種;以ACh為例,當它們作用於神經-肌接頭處時,終板膜上有同它們作特異結合的化學門控通道;但當ACh作用於心肌或內臟平滑肌時,遇到的卻是受體-G-蛋白-第二信使系統(受體稱為M-型毒蕈礆型受體)。由此可見,同一種刺激信號通過何種跨膜信號傳遞系統起作用,關鍵因素在於靶細胞膜上具有何種感受結構;近年還發現,即便是M-型ACh受體,也可再區分出許多種亞型,有的亞型以cAMP為第二信使,有的以IP3和DG為第二信使。不同細胞甚或同一細胞的膜上具有對應於同一化學信號的不同受體型或其亞型,在跨膜信號傳遞中並不少見。近年來發現基本嗅覺刺激(大約是7種)全都是通過嗅上皮中不同的膜受體-第二信使系統起作用的,但在4種基本味覺刺激中,只有咸和酸刺激是通過細胞上相應的化學門控上通道起作用的,甜味物質是通過受體-第二信使系統起作用的,而苦味物質則因物質分子不同而分別通過通道和受體兩種途徑起作用。第三,跨膜信號傳遞的方式雖然相對地較少,但也不一定只限於上述兩種。近年來有一些特殊的化學信號影響其靶細胞的方式受到廣泛的重視,很可能成為跨膜信號傳遞的一種新類型;這就是發現胰島素等一些肽類激素和其他與機體發育、生長、修復、增生、甚至細胞癌變有關的因子,如神經生長因子、表皮生長因子、血小板源生長因子、纖維母細胞生長因子、以及與細胞生成有關的集落刺激因子等,都是通過靶細胞表面一類稱為酪氨酸激酶受體(tyrosine kinase riceptor)的蛋白質起作用的,這類受體結構簡單,只有一個跨膜α-螺旋,當位於膜外側的較長的肽鏈部分同特定的化學信號結合後,可以直接引起受體肽鏈的膜內段激活,使之具有磷酸激酶活性,通過使自身肽鏈和膜內蛋白質底物中的酪氨酸殘基發生磷酸化,因而產生細胞內效應。這方面的新資料正在積累之中。
癌基因和跨膜信號傳遞近年發現與上述跨膜信號傳遞有關的一些蛋白質,如受體、G-蛋白、各種生長刺激因子和營養因子、以及各種蛋白激酶等,它們在細胞內的生物合成,是由人正常染色體中被稱為細胞原癌基因(cellular proto-oncogene,進行表達時稱細胞癌基因)的一類基因所編碼和表達生成的。這些基因所以被稱為原癌基因,是因為它們的礆基排列順序同一些(近100種)能在動物引起腫瘤的病毒DNA(或稱病毒癌基因,viral oncogene)的礆基排列順序相一致。關於細胞癌基因與人類腫瘤發生的關係目前尚不清楚,但它們的正常表達產物,卻是人體無時無刻不在進行著的各種跨膜信號傳遞過程所必需的。試設想,如果由於遺傳和變異等原因使細胞不能合成結構和功能正常的G-蛋白,對人體將會有何等廣泛而重要的影響!另外,在細胞原癌基因中,有一類可被胞漿中產生的第二信使等物質所激活,生成某種蛋白質;但它們在胞漿中生成後,一般又進入核內,進而誘導另一些基因進行表達。這類癌基因從激活到蛋白質生成,比一般基因表達為快,稱為快速基因(或即早基因),而它們生成的蛋白質的作用則是激活另一些(可能在細胞功能活動中更重要)基因的表達,故快速基因的表達產物可稱為轉錄調節因子或第三信使。所以稱為第三信使是因為它們由第二信使類物質的作用而生成,而它們自身的作用又引起新的基因表達,生成一些可能對細胞結構和功能有較長遠影響的蛋白質。這樣外來信號(第一信使)不僅通過第二信使的合成在胞漿中引起一些即時反應,還可能通過第三信使引起細胞功能和結構長時間的適應性改變。
| 關於「生理學/由膜的特異受體蛋白質、G-蛋白和膜的效應器酶組成的跨膜信號傳遞系統」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |