生物化學與分子生物學/成熟紅細胞的代謝特點
| 醫學電子書 >> 《生物化學與分子生物學》 >> 血漿蛋白與凝血 >> 紅細胞的代謝 >> 成熟紅細胞的代謝特點 |
| 生物化學與分子生物學 |
|
|
成熟紅細胞不僅無細胞核,而且也無粒線體、核蛋白體等細胞器,不能進行核酸和蛋白質的生物合成,也不能進行有氧氧化,不能利用脂肪酸。血糖是其唯一的能源。紅細胞攝取葡萄糖屬於易化擴散,不依賴胰島素。成熟紅細胞保留的代謝通路主要是葡萄糖的酵解和磷酸戊糖通路以及2.3一二磷酸甘油酸支路(2,3-biphosphoglycerate,2.3桞PG)。通過這些代謝提供能量和還原力(NADH,NADPH)以及一些重要的代謝物(2,3桞PG),對維持成熟紅細胞在循環中約120的生命過程及正常生理功能均有重要作用。
(一)糖酵解
循環血液中的紅細胞每天消耗約30g葡萄糖,其中90~95%經糖酵解被利用。一分子葡萄糖經酵解可產生2分子ATP。紅細胞中生成的ATP主要用於維持紅細胞膜上的離子泵(鈉泵、鈣泵),以保持紅細胞的離子平衡;維持細胞膜可塑性;谷胱甘肽合成及核苷酸的補救合成等。缺乏ATP則紅細胞膜內外離子平衡失調,紅細胞內Na+進入多於K+排出、Ca++進入增多,紅細胞因吸入過多水分而膨大成球狀甚至破裂。同時由於ATP缺乏,可使紅細胞膜可塑性下降,硬度增高,易被脾臟破壞,造成溶血。
紅細胞無氧酵解中生成的NADH+H+是高鐵血紅蛋白還原酶的輔助因子,此酶催化高鐵血紅蛋白還原為有載氧功能的血紅蛋白。
(二)2,3-二磷酸甘油酸(2,3-BPG)支路
在糖無氧酵解通路中,1,3-二磷酸甘油酸(1,3-BPG)有15~50%在二磷酸甘油酸變位酶催化下生成2,3-BPG,後者再經2,3-BPG磷酸酶催化生成3磷酸甘油酸。經此2,3-BPG的側支循環稱2,3-BPG支路(圖10-19)。
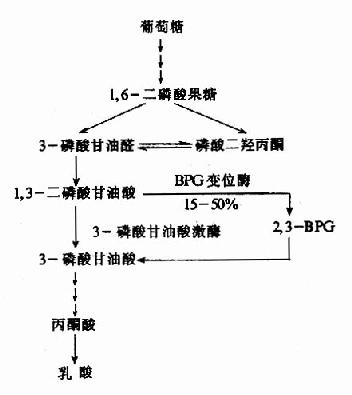
圖10-19 2,3-BPG支路
紅細胞中2,3-BPG磷酸酶活性遠低於BPG變位酶,使2,3-BPG的生成大於分解,因而紅細胞中2,3-BPG的濃度處於有機磷酸酯的巔峰,較糖酵解其它中間產物的有機磷酸酯高出數+甚至數百倍(表10-4)。
表10-4 紅細胞中各種糖酵解中間產物的濃度(微克分子/升紅細胞)
| 糖酵解中間產物 | 動脈血 | 靜脈血 |
| 6-磷酸葡萄糖 | 30.0 | 24.8 |
| 6-磷酸果糖 | 9.3 | 3.3 |
| 1,6二磷酸果糖 | 0.8 | 1.3 |
| 磷酸丙糖 | 4.5 | 5.0 |
| 3-磷酸甘油酸 | 19.2 | 16.5 |
| 2-磷酸甘油酸 | 5.0 | 1.9 |
| 磷酸烯醇式丙酮酸 | 10.8 | 6.6 |
| 丙酮酸 | 87.5 | 143.2 |
| 2,3-DPG | 3400 | 4940 |
2,3-BPG能特異地與去氧血紅蛋白(deoxy Hb)結合,2,3-BPG進入血紅蛋白α2β2四聚體中心空隙兩個β亞基之間,借其分子中所帶5個負電荷與兩個β亞基的帶正帶胺基酸殘基以鹽鍵及氫鍵結合,使兩個β亞基保持分開的狀態,即促使血紅蛋白由緊密態向松馳態轉換,從而減低血紅蛋白對氧的親和力(圖10-20)。
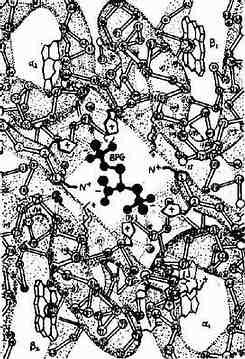
圖10-20 BPG與Hb的作用示意圖
當紅細胞內2,3-BPG濃度升高時有利於HbO2放氧,而2,3-BPG濃度下降則有利於Hb與氧結合。BPG變位酶及2,3-BPG磷酸酶受pH值調節。在肺泡毛細血管血液pH高,BPG變位酶受抑制而2,3-BPG磷酸酶活性強。使紅細胞內2,3-BPG的濃度降低,有利於Hb與O2結合。
反之,在外周組織毛細血管中,血液pH下降,2,3-BPG的濃度升高,則利於HbO2放氧,藉此調節氧的運輸和利用,具有重要生理意義。但2,3-BPG的生成是以減少一個ATP的生成為代價的。
(三)磷酸戊糖通路
紅細胞內利用葡萄糖的5~10%通過磷酸戊糖通路代謝,為紅細胞提供另一種還原力(NADPH),NADPH在紅細胞氧化還原系統中發揮重要作用,具有保護膜蛋白、血紅蛋白及酶蛋白的巰基不被氧化,還原高鐵血紅蛋白等多種功能。
1.GSH的主要生理功能是對抗氧化劑對巰基的氧化。細胞內可自發生成少量超氧陰離子(O-2),同時感染時的白細胞吞噬作用亦可產生O-2,可被超氧化物歧化酶(superoxidedismufase SOD),催化生成過氧化氫(H2O2)。
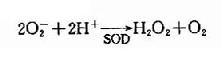
而GSH在谷胱甘肽過氧化酶作用下將H2O2還原為H2O,GSH自身被氧化為氧化型谷胱甘肽(GSSG)。後者在谷胱甘肽還原酶催化下,由NADPH+H+供氫重新還原為GSH。(圖10-21)。
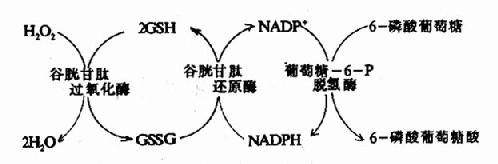
圖10-21 谷胱甘肽的氧化與還原
催化NADPH生成的關鍵酶為葡萄糖-6-磷酸脫氫酶。此酶缺陷的病人一般情況下無症状,但有外界因素(如進食某種蠶豆)影響,即引起溶血。因吃蠶豆可誘導發病,故這種病又稱蠶豆病。
2.高鐵血紅蛋白(methemoglobin MHb)的還原:由於各種氧化作用,紅細胞內經常有少量MHb產生,而由於紅細胞內有一系列酶促及非酶促的MHb還原系統(表10?),故正常紅細胞中MHb只佔1-2%。
表10-5 紅細胞中MHb還原系統
| 還原系統 | 佔總還原能力的百分比(%) |
| 酶促還原系統 | |
| NADH脫氫酶Ⅰ | 61 |
| NADH脫氫酶Ⅱ | 5 |
| NADH脫氫酶 | 6 |
| 非酶促還原系統 | |
| 抗壞血酸 | 16 |
| 谷胱甘肽 | 12 |
由表10-5可知,催化MHb還原的主要是NADH-脫氫酶,輔酶為NADH+H+。NADPH脫氫酶,(以NADPH+H+為輔酶)也參與MHb還作,但作用較小。除此之外,抗壞血酸和GSH可直接還原MHb,而氧化型抗壞血酸和GSSG的還原作用最終需NADPH+H+供氫。
| 關於「生物化學與分子生物學/成熟紅細胞的代謝特點」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |