醫學微生物學/細菌的突變
| 醫學電子書 >> 《醫學微生物學》 >> 細菌的遺傳與變異 >> 細菌的突變 |
| 醫學微生物學 |
|
|
遺傳型變異中常見的一種為突變(Mutation),即細菌的基因結構發生偶然的改變。一般突變會導致所編碼蛋白質的改變,從而使細菌出現新的特性或失去原有的某些特性。細菌的自然突變率與其生物的自然突變相同,每106~108次細胞分裂發生一次。由於細菌每20~30分鐘分裂一代,故突變株相對較多。當突變發生在DNA一對或少數幾對鹼基引起改變時,稱為點突變。這類突變涉及鹼基對的置換、增加或缺失。另一種類型的突變涉及大段DNA的改變如插入或缺失幾百個鹼基對,稱為染色體畸變。在細菌中點突變較多見,但在大腸桿菌的染色體中曾發現有800~1400鹼基對的插入,稱為插入順序(Insertion sequences)。
一、突變與選擇
由於細菌的突變所發生的表型變異必須在一定外界環境下才表現出來,因此對外界環境在突變中的作用曾發生過爭議。曾有學者認為細菌與其他生物一樣,需經過突變與外界環境條件選擇而出現突變株;然而也有學者認為細菌生長繁殖迅速,外界環境可直接作用誘導出變異株。這一爭論直到1943年Luria與Delbruck進行了有名的變異試驗(Fluctuation test)後才初步獲得解決。變異試驗是根據統計學原理設計的(圖5-1)。將一瓶對大腸桿菌噬菌體T1敏感的細菌培養後,稀釋至1,000細菌/ml,分別分至兩套試管系列。一套僅將細菌接種至一支大管培養基中,經一段時間培養後,分別接種於數個平板。在平板中加入噬菌體,(如果出現耐噬菌體裂解的突變細菌株則在該平板上應出現菌落)以篩選耐噬菌體的突變株菌落。另一套試驗為將細菌分別接種於數支小試管的培養基,經過一段時間後自每支試管吸取菌液各塗布接種一個加入噬菌體的平板,然後計數發生的突變耐噬菌體菌株菌落數。如果出現耐噬菌體菌株是因接觸噬體而誘導產生的,兩套平權上出現的耐噬菌體菌株菌落數。如果出耐噬菌體菌株是接觸噬體而誘導產生的,兩套平權上出現的噬菌體菌落數應大體相等。如果系細菌自發突變則兩套平板上的耐噬菌體菌落數應有顯著不同,因為在後一情況下,細菌分別在各支試管中各自在生長繁殖周期或早或遲可發生突變。突變發生早的突變株繁殖後出現的子代數必然多於突變株發生晚者,因此各平板上的菌落有的很多,有的則缺如。實驗結果證明兩套平板上耐噬菌體菌落數差異很大,從萬里證實細菌已自身發生為,而外界是條件僅起選擇作用。
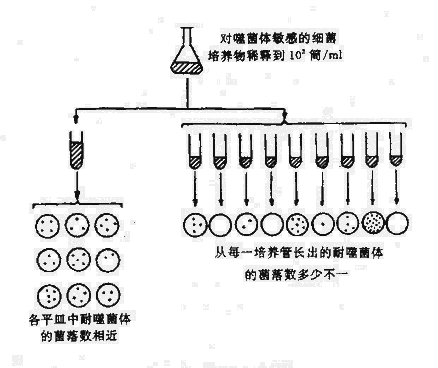
圖5-1 細菌的變異試驗
然而,仍有學者對上述實驗的統計學意義特異議。1952年Lederberg設計了影印培養(Replica plating)(見圖5-2),這一試驗證明細菌耐藥突變不是由抗菌藥物誘導師產生,而在未接觸抗菌藥物前已產生耐藥突變。其方法為將對鏈黴素敏感的大腸桿菌大量接種於營養瓊脂平板上,培養後細菌在培養基上均勻的長出菌苔層,然後用滅菌的絲絨復蓋在一塊與平皿同樣大小的木塊上輕輕影印,使細菌菌落全部轉移到絲絨面上。另取含有鏈黴素的瓊脂培養平板,將絲絨面再印在其上,從而獲得與原始平板完全相同的複製平板。將此平板培養,耐藥的菌落出現後,通過比較,可從原始平板的相應部位刮取菌苔轉種至液體培養基中,增菌後,重複製備原始平板與影印平板。經過數次重複後,則可獲得一株完全沒有接觸過鏈黴素的高度耐鏈毒素的突變株,表現為原始平板上全部菌落與含鏈黴素平板上的菌落吻合。這一實驗雄辯地證明了鏈黴素僅起選擇作用。
二、基因突變類型
在明確了細菌培養環境條件僅起選擇作用後,學者們採用了一些選擇方法以獲得突變株,這些突變株的應用對研究遺傳變異及開展有基因工程都很有幫助。大從數利用的突變型均為大腸桿菌或鼠傷寒沙門氏菌,因這兩種細菌營養要求不高,僅需鹽、微量金屬離子及有限的氮、碳來源即可生長繁殖。未發生突變的菌株稱為野生型,而發生突變的菌株則根據選擇條件可分為:
1.營養缺陷型:突變後因某種酶的缺失需要額外添加某種營養成份方能生長繁殖者。一般用「+」代表能利用自然存在的某種成份或能合成某種成份的中間體,而「—代表不能合成該成份的菌株。如his-則代表組氨酸缺陷型,需在培養基中加入組氨酸。
2.抗性突變型:一般以S代表對化學藥物或抗生素敏感,r代表有抵抗力。如str8代表該菌株對鏈黴素敏感,在有鏈黴素存在時不能生長。這類突變型最易獲得,應用亦廣。
3.發酵陰性突變型:突變後失去發酵某種糖的能力但仍能利用其他糖做為碳源。這是由於突變後失去能分解該糖的酶。由於乳糖發酵可用指示劑根據pH改變而顯示,故可Lac-(乳糖發酵陰性)突變株作為研究工具。
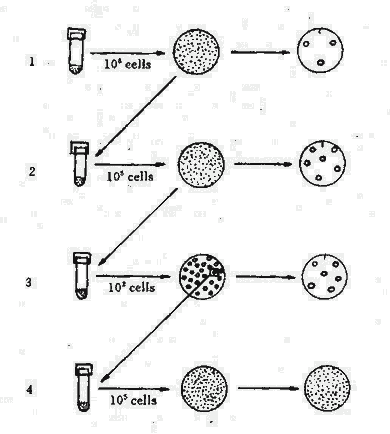
圖5-2 細菌影印培養試驗
| 1.從普通培養基上將皿上長的細菌,用絲絨影印至含鏈黴素培養基上,耐菌細菌不多。 |
| 2.從不接觸鏈黴素的平皿上,按影印顯示耐鏈黴素菌落區挑取細菌增菌。 |
| 3.將102細菌接種普通培養基上再作影印,再按耐鏈黴素平皿上相應菌落處接種增菌。 |
| 4.未接觸鏈黴素的平皿上,全部細菌均耐鏈黴素。 |
4.條件致死性突變型:在某一條件下具有致死效應,突變株不能生長,但在另一沒有致死效應的條件下仍可生長。最常用者為溫度敏感性突變型。它們在親代能生長的溫度範圍內特別是較高濕度(42℃)不能生長,但在較低溫度(25℃)則能生長。這種菌株稱為ts株。其發生的原因是某些酶的肽鏈結構發生改變後,降低了酶的抗熱性。因此在較高溫度下不能生存。這種用溫度篩選突變株的方法比較簡便,應用較多。
三、基因突變的分子生物學基礎
細菌的基因結構發生改變的機制包括:
1.鹼基置換(Substitution):包括兩種類型:轉換(Transition)是由嘌呤置換嘌呤或嘧啶置換嘧啶。顛換(Transversion)是指嘌呤置換嘧啶或嘧啶置換嘌呤。如鹼基置換髮生於編碼多肽的區,則因可影響密碼子而使轉錄、翻譯遺傳信息發生變化,因此可以出現一種胺基酸取代原有的某一種胺基酸。也可能出現了終止密碼而使多肽鏈合成中斷,不能形成原有的蛋白質而完全失去某種生物學活性。
2.鹼基的減少、增加與倒置:三種情況都可造成對密碼的錯誤閱讀。如DNA原有鹼基順序為AAG,GAA,CGC,TGA,如失去第一個A,則成為AGG,AAC,GCT,GA,使原來編碼押肽由亮一組一丙一蘇氨酸改為半胱-亮-精-亮氨酸。這種突變導致的是密碼意義的錯誤,稱為移碼突變。移碼突變的影響範圍自突變點起直到末端整條結構基因的轉錄與翻譯,引起基因產物的變化比較嚴重,對生物活性的影響也較顯著。
3.鹼基的互變異構:四種鹼基中的任何一種均可發生互變異構,在作為模板時可引起互補鹼基的改變。如當胸腺嘧啶以正常形式(即酮基型)為模板時,配對的互補鹼基為腺嘌呤;當前者變為烯醇型結構時,通過氫鍵配對的鹼基可變為鳥嘌呤。(圖5-3)
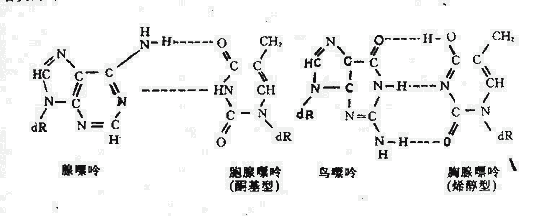
圖5-3 鹼基的互變異構
細菌自發突變的發生原因可能是宇宙間普遍存在的短波輻射、熱及自然界存在的一些具有致突變作用的物質。人工應用理化因素可誘發突變者稱為誘變劑。化學誘變劑包括核苷酸鹼基的類似物如分子結構類似胸腺嘧啶的5-溴尿嘧啶。烷化劑可改變鹼基的化學結構也是誘變劑。吖啶類染料可螯合入DNA的鹼基對之間,引起DNA在複製過程中出現鹼基對的插入或缺失。紫外線與X線是常用的物理誘變劑。紫外線可使鄰近的胸原嘧啶構成雙體,引起DNA結構的變化而致突變。
| 關於「醫學微生物學/細菌的突變」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |