臨床生物化學/LDL受體
| 醫學電子書 >> 《臨床生物化學》 >> 血漿蛋白及其代謝紊亂 >> 脂蛋白受體 >> LDL受體 |
| 臨床生物化學 |
|
|
最先從牛腎上腺分離出LDL受體,以後又分離了編碼牛LDL受體羥基末端1/3胺基酸的cDNA,並初步闡明了牛LDL受體的cDNA,並且推導出人LDL受體的胺基酸序列。
(一)LDL受體結構
LDL受體是一種多功能蛋白,由836個胺基酸殘基組成36面體結構蛋白,分子量約115ku,由五種不同的區域構成,各區域有其獨特的功能,見圖4-6。
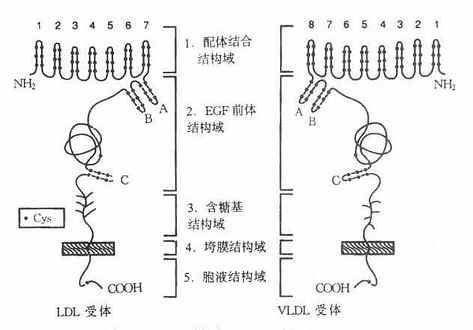
圖4-6 LDL受體與VLDL受體結構示意圖
1.配體結合結構域配體結合結構域由292個胺基酸殘基組成,其中共有47個半胱氨酸(Cys)。含有七個由40個殘基組成的與補體Cb和Cq類似的重複序列,每個重複系列中有6個半胱氨酸殘基,所有42個半胱氨酸殘基均已構成二硫鍵,重複序列2,3,6,7是結合LDL所必需,其中任何一種發生突變,均使受體喪失結合LDL的能力。重複序列5則與結合β-VLDL有關,若該序列突變時,受體結合β-VLDL的能力喪失60%。該受體不僅能結合LDL,還能結合VLDL、β-VLDL和VLDL殘粒,它不僅能識別ApoB100,也可識別含ApoE的脂蛋白。ApoE、B100為LDL受體的配體,因此,LDL受體又稱為ApoB100E受體。
2.EGF前體結構域該域約由400個胺基酸殘基組成的肽段,有五個重複序列,每個重複序列包括25個胺基酸殘基。EGF前體結構域與小鼠上皮細胞生長因子(epidermalgrowthfactor,EGF)前體有同源性,這一區域因此而得名。體外實驗證實,這個區域的肽段,屬於細胞膜外結構蛋白,起著支撐作用。
3.糖基結構域由58個胺基酸殘基組成,是緊靠細胞膜面的肽段,有18個絲氨酸或蘇氨酸,構成O-連接糖鏈,對LDL受體也有支撐作用。
4.跨膜結構由22個胺基酸殘基組成,富含疏水胺基酸殘基,屬於跨膜蛋白,起著固繫於細胞膜中的「拋錨」作用。這個區域若有缺陷則影響受體的細胞外分泌。
5.胞液結構域位於細胞膜的胞質側,由50個胺基酸殘基組成,C-末端位於胞質並「深埋」於胞質之中。
(二)LDL受體基因結構及功能
1.受體親和性含ApoB100的脂蛋白可以與LDL受體以高親和力結合,腸道分泌的ApoB48不是LDL受體的配體,所以肝臟不能清除完整的CM。
2.基因結構人LDL受體基因長度45ku,由18個外顯子和17個內含子組成。
3.LDL受體途徑LDL受體廣泛分布於肝、動脈壁平滑肌細胞、腎上腺皮質細胞、血管內皮細胞、淋巴細胞、單核細胞和巨噬細胞,各組織或細胞分布的LDL受體活性差別很大。
LDL或其他含ApoB100、E的脂蛋白如VLDL、β-VLDL均可與LDL受體結合,內吞入細胞使其獲得脂類,主要是膽固醇,這種代謝過程稱為LDL受體途徑(LDLreceptorpathway)。該途徑依賴於LDL受體介導的細胞膜吞飲作用完成,如圖4-7所示。當血漿中LDL與細胞膜上有被區域(coatedregion)的LDL受體結合(第1步),使其出現有被小窩(coatedpit)(第2步),並從膜上分離形成有被小泡(coatedvesicles)(第3步),其上的網格蛋白(clathrin)解聚脫落,再結合到膜上(第4步),其內的pH值降低,使受體與LDL解離(第5步),LDL受體重新回到膜上進行下一次循環(第6、7步)。有被小泡與溶酶體融合後,LDL經溶酶作用,膽固醇酯水解成游離膽固醇和脂肪酸,甘油三酯水解成脂肪酸,載脂蛋白B100水解成胺基酸。LDL被溶酶體水解形成的游離膽固醇再進入胞質的代謝庫,供細胞膜等膜結構利用。胞內游離膽固醇在調節細胞膽固醇代謝上具有重要作用;若胞內濃度升高,可能出現下述種情況:①抑制HMGCoA還原酶,以減少自身的膽固醇合成;②抑制LDL受體基因的表達,減少LDL受體的合成,從而減少LDL的攝取,這種LDL受體減少的調節過程稱為下調(downregulation);③激活內質網脂醯基CoA膽固醇醯轉移酶(Acyl-CoAcholesterolacyltransferase,ACAT),使游離膽固醇在胞質內酯化成膽固醇酯貯存,以供細胞的需要。經上述三方面的變化,用以控制細胞內膽固醇含量處於正常動態平衡狀態。血漿中膽固醇主要存在於LDL中,而65%-70%的LDL是依賴肝細胞的LDL受體清除。肝的LDL受體還影響LDL的合成速率及VLDL代謝。曾經認為人VLDL幾乎全部在血循環中轉變為LDL,LDL再被肝外組織攝取。現在經大鼠和兔實驗研究表明,僅有15%以下轉變為LDL,人則是小於50%的VLDL轉變為LDL,大部分VLDL是以VLDL或VLDL殘粒的形成被肝攝取。VLDL殘粒與肝受體的親和力比VLDL大很多。所以VLDL殘粒被肝清除的速率比VLDL快。VLDL殘粒大部分被肝清除,一小部分在肝脂酶作用下水解除去甘油三酯而轉變成LDL。LDL受體還在乳糜微粒代謝中起一定作用。乳糜微粒中的ApoB48不能識別ApoB100E受體,所以肝不能清除完整的乳糜微粒。CM中雖有少量ApoE,因含有豐富的ApoC,可掩蓋ApoE,而阻礙其與肝的ApoB、E受體結合,血液中乳糜微粒被脂蛋白脂肪酶水解去除其大部分甘油三酯核心後,同時喪失部分ApoC、A,生成乳糜微粒殘粒後除去了阻礙ApoE與受體結合的因素,其殘粒可迅速被肝清除,約有一半是通過LDL受體,另一半通過LDL受體相關蛋白代謝,其半壽期短。
總之,LDL受體主要功能是通過攝取Ch進入細胞內,用於細胞增殖和固醇類激素及膽汁酸鹽的合成等。
| 關於「臨床生物化學/LDL受體」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |