生理學/視桿細胞的感光換能機制
| 醫學電子書 >> 《生理學》 >> 感覺器官 >> 視覺器官 >> 視桿細胞的感光換能機制 |
| 生理學 |
|
|
|
從上世紀末開始,有人就從視網膜中提取出了一定純度的感光色素即視紫紅質,它在暗處呈紅色;實驗中還可以證明,提取出來的這種感光色素對不同波長光線的吸收光譜,基本上和晚光覺對光譜不同部分的敏感性曲線相一致(圖9-7)。這一事實十分重要,因為既然光線對某種感光色素的光化學作用的強度正好與這些光線所引起的視覺的強度相一致,那就是提示前者可能是後者的基礎。
(一)視紫紅質的光化學反應及其代謝
視紫紅質的分子量約為27-28kd,是一種與結合蛋白質,由一分子稱為視蛋白(opsin)的蛋白質和一分子稱為視黃醛(retnal)的生色基團所組成。視蛋白的肽鏈序列已搞清,它的肽鏈中有7段穿越所在膜結構、主要由疏水性胺基酸組成的α-螺旋區段,同一般的細胞膜受體具有類似的結構。視黃醛由維生素A變來,後者是一種不飽和醇,在體內一種酶的作用下可氧化成視黃醛。提純的視紫紅質在溶液中對500nm波長的光線吸收能力最強,這與人眼在弱光條件下對光就業上藍綠光區域(相當於500nm波長附近)感覺最明亮(不是感到了藍綠色)的事實相一致(圖9-7),說明人在暗視覺與視桿細胞中所含視紫紅質的光化學反應有直接的關係。
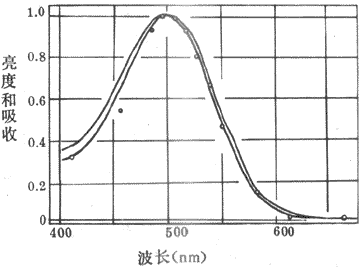
圖9-7弱光條件下人眼所感到的光譜亮度曲線和實驗條件下
視紫紅質對光譜不同部分的吸收曲線視覺中最明亮的區域和視紫紅制裁
吸收能力最強的部分都在500nm的波長附近
視紫紅質在光照時迅速分解為視蛋白和視黃醛,這是一個階段的反應。目前認為,分解的出現首先是由於視黃醛分子在光照時發生了分子構象的改變,即它在視紫紅質分子中本來呈11-順型(一種較為彎曲的構象),但在光照時變為全反型(一種較為直的分子構象)。視黃醛分子構象的這種改變,將導致視蛋白分子構象也發生改變,經過較複雜的信號傳遞系統的活動,誘發視桿細胞出現感受器電位。據計算,一個光量子被視紫紅質吸收,就足以使視黃醛分子結構發生改變,導致視紫紅質最後分解為視蛋白和視黃醛。視紫紅質分解的某些階段伴有能量的釋放,但這看來不是誘發感受器電位的直接原因。
在亮處分解的視紫紅質,在暗處又可重新合成,亦即它是一個可逆反應,其反應的平衡點決定於光照的強度。視紫紅質再合成的第一步,是全反型的視黃醛變為11-順型的視黃醛,很快再同視蛋白結合。此外,貯存在視網膜的色素細胞層中的維生素A也是全反型的,它們也可在耗能的情況下變成11-順型的,進入視桿細胞,然後再氧化成11-順型的視黃醛,參與視紫紅質的合成補充;但這個過程進行的速度較慢,不是促進視紫紅制裁再合成的即時因素。人在暗處視物時,實際是既有視紫紅質的分解,又有它的合成,這是人在暗光處能不斷視物的基礎;光線愈暗,全盛過程愈超過分解過程,視網膜中處於合成狀態的視紫紅質數量也愈高,這也使視網膜對弱光愈敏感;相反,人在亮光處時,視紫紅質的分解增強,合成過程甚弱,這就使視網膜中有較多的視紫紅質處於分解狀態,使視桿細胞幾乎失去了感受光刺激的能力;事實上,人的視覺在亮光處是靠另一種對光刺激較不敏感的感光系統即視錐來完成的,後一系統在弱光時不足以被刺激,而在強光系統下視桿細胞中的視紫紅質較多地處於分解狀態時,視錐系統就代之而成為強光刺激的感受系統。在視紫紅質和再合成的過程中,有一部分視黃醛被消耗,這最終要靠由食物進入血液循環(相當部分貯存於肝)中的維生素A來補充。長期攝入維生素A不足,將會影響人在暗光處的視力,引起夜盲症。
(二)視桿細胞外段的超威結構和感受器電位的產生
感光細胞的外段是進行光-電轉換的關鍵部位。視桿細胞外段具有特殊的超威結構,如圖9-8所示。在外段部分,膜內的細胞漿甚少,絕大部分為一些整齊的重疊成層的圓盤狀結構所佔據,這圓盤稱為視盤。每一個視盤是一個扁平的囊狀物,囊膜的結構和細胞膜類似,具有一般的脂質雙分子層結構,但其中鑲嵌著的蛋白質絕大部分是視紫紅質,亦即視桿細胞所含的視紫紅質實際上幾乎全部集中在視盤膜中。視盤的數目在不同動物的視桿細胞中相差很大,人的每個視桿細胞外段中它們的數目近千;每一個視盤所含的視紫紅質分子約有100萬個。這樣的結構顯然有利於使進入視網膜的光量子有更大的機會在外段中碰到視紫紅質分子。
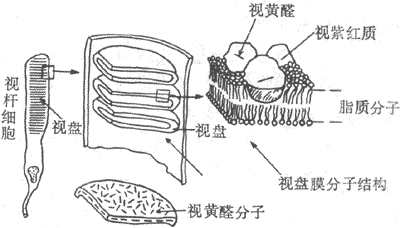
圖9-8 視桿細胞外段的超微結構示意圖
有人用細胞內微電極技術,研究了視桿細胞外段內外的電位差在光照前後的變化,結果發現在視網膜未經照射時,視桿細胞的靜息電位只有-30?/FONT>-40mV,比一般細胞小得多。經分析表明,這是由於外段膜在無光照時,就有相當數量的Na+通道處於開放狀態並有持續的Na+內流所造成,而內段膜有Na+泵的連續活動將Na+移出膜外,這樣就維持了膜內外的Na+平衡。當視網膜受到光照時,可看到外段膜兩側電位短暫地向超極化的方向變化,由此可見,外段膜同一般的細胞膜不一致,它是在暗處或無光照時處於去極化狀態,而在受到光刺激時,跨膜電痊反而向超極化方向變化,因此視桿細胞的感受器電位(視錐細胞也一樣),表現為一種超極化型的慢電位,這在所有被研究過的發生器或感受器電位中是特殊的,它們一般都表現為膜的暫時去極化。
光子的吸收引起外段膜出現超極化電反應的機制已基本搞清,這就是光量子被作為受體的視紫紅質吸收後引起視蛋白分子的變構,又激海參了視盤膜中一種稱為傳遞蛋白(transducin)Ct的中介物,後者在結構上屬於G-蛋白家庭的一員,它激活的結果是進而激活附近的磷酸二酯酶,於是使外段部分胞漿中的cGMP大量分解,而胞漿中cGMP的分解,就使未受光刺激時結合於外段膜的cGMP由也膜解離而被分解,而cGMP在膜上的存在正是這膜中存在的化學門控式Na+通道開放的條件,膜上cGMP減少,Na+通道開放減少,於是光照的結果出現了我們記錄到的超極化型感受器電位。據估計,一個視紫紅質被激活時,可使約500個傳遞蛋白被激活;雖然傳遞蛋白激活磷酸二酯酶是1對1的,但一個激活了的磷酸二酯酶在一秒鐘內大約可使4千多個cGMP分子降解。由於酶系統的這種生物放大作用,就可以說明1個光量子的作用何以能在外段膜上引起大量化學門控式Na+通道的關閉,引起一個足以為人的視覺系統所感知的超極化型電變化。
視桿細胞外段和整個視桿細胞都沒有產生動作電位的能力,由光刺激在外段膜上引起的感受器電位只能以電緊張性的擴布到達它的終足部分,影響終點(相當於軸突末稍)外的遞質釋放。
| 關於「生理學/視桿細胞的感光換能機制」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |