生物化學與分子生物學/RNA轉錄作用
| 醫學電子書 >> 《生物化學與分子生物學》 >> RNA的生物合成 >> RNA轉錄作用 |
| 生物化學與分子生物學 |
|
|
|
RNA的轉錄合成從化學角度來講類似於DNA的複製,多核苷酸鏈的合成都是以5』→3』的方向,在3』-OH末端與加入的核苷酸磷酸二酯鍵,但是,由於複製和轉錄的目的不同,轉錄又具有其特點:(1)對於一個基因組來說,轉錄只發生在一部分基因,而且每個基因的轉錄都受到相對獨立的控制(圖17-2);(2)轉錄是不對稱的。(3)轉錄時不需要引子,而且RNA鏈的合成是連續的。
目錄 |
一、依賴DNA的RNA聚合酶
真核和原核細胞內都存在有DDRP,迄今發現的DDRP的有以下特點:①以DNA為模板;在DNA的兩條多苷酸鏈中只有其中一條鏈作為模板,這條鏈叫做模板鏈(templatestrand),又叫做無意義鏈。DNA雙鏈中另一條不做為模板的鏈叫做編碼鏈,又叫做有意義鏈,編碼鏈的的序列與轉錄本RNA的序列相同,只是在編碼鏈上的T在轉錄本RNA為U,由於RNA的轉錄合成是以DNA的一條鏈為模板而進行的,所以這種轉錄方式又叫做不對稱轉錄。②都以四種三磷酸核苷的底物,原料;③都遵循DNA與RNA之間的鹼基配對原由,A=U,T=A,C=G,合成與模板DNA序列互補的RNA鏈。④RNA鏈的延長方向是5』→3』的連續合成。⑤需要Mg2+或Mn2+離子。⑥並不需要引子。RNA聚合酶缺乏3』→5』外切酶活性,所以沒有校正功能。
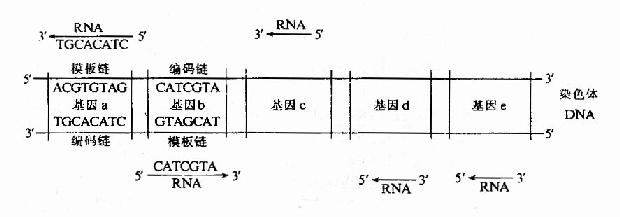
圖17-2 細菌染色體上幾個基因轉錄的方向及所用模板
RNA聚合酶催化下列反應:
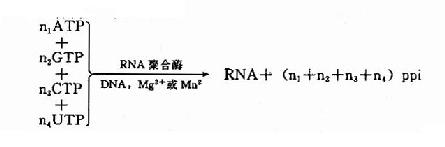
大腸桿菌RNA聚合酶的研究得比較透徹的,這是一個分子量達50多萬,全酶由五咱亞基組成,去掉δ亞基的部分稱為核心酶,核心酶本身就能催化苷酸間磷酸二酸鍵形成。利福平和利福黴素能結合在β亞基上而對此酶發生強烈的抑制作用。β亞基似乎是酶和核苷酸底物結合的部位。細胞內轉錄是在DNA特定的起始點上開始的,δ亞基的功能是辨認轉錄起始點的。β』亞基是酶與DNA模板結合的主要成分。α亞基可能與轉錄基因的類型和種類有關。
表17-1 大腸桿菌RNA聚合酶
| 亞單位 | 分子量 | 亞單位數目 | 功能 |
| α β β δ |
36512 150618 155613 70263 |
2 1 1 1 |
決定哪此基因被轉錄 與轉錄全過程有關 結合DNA模板 辨認起始點 |
真核生物中已發現有四種RNA聚合酶,分別稱為RNA聚合酶Ⅰ、Ⅱ、Ⅲ和粒線體RNA聚合酶,分子量大致都在50萬道爾頓左右,它們專一性地轉錄不同的基因,因此由它們催化的轉錄產物也各不相同。RNA聚Ⅰ合成RNA的活性最顯著,它位於核仁中,負責轉錄編碼rRNA的基因。而細胞內絕大部分RNA是rRNA是RNA。RNA聚合酶Ⅱ,位於核質中,負責核內不勻一RNA的合成,而hnRNA是mRNA的前體。RNA聚合酶Ⅲ負責合成tRNA和許多小的核內RNAs。鵝膏蕈鹼是真核生物RNA聚合酶特異性抑制劑,三種真核生物RNA聚合酶對鵝膏蕈鹼的反應不同,見表17-2,原核生物靠RNA聚合酶就可完成從起始、延長、終止的轉錄全過程,真核生物轉錄除RNA聚合酶外還需另一此叫做轉錄因子的蛋白質分子參與轉錄的全過程,見後敘。
表17-2 真核生物的RNA聚合酶
| 種類 | 分布 | 合成的RNA類型 | 對α-鵝膏蕈鹼的敏感性 |
| Ⅰ Ⅱ Ⅲ Mt |
核仁 核質 核質 粒線體 |
rRNa hnRNA tRNA,5sRNA 粒線體RNAs |
不敏感 低濃度敏感 高濃度敏感 不敏感 |
二、RNA的轉錄過程
轉錄的主要過程見圖17-3,為研究和敘述方便,可將RNA合成分為識別與起始、延長和終止三個階段。
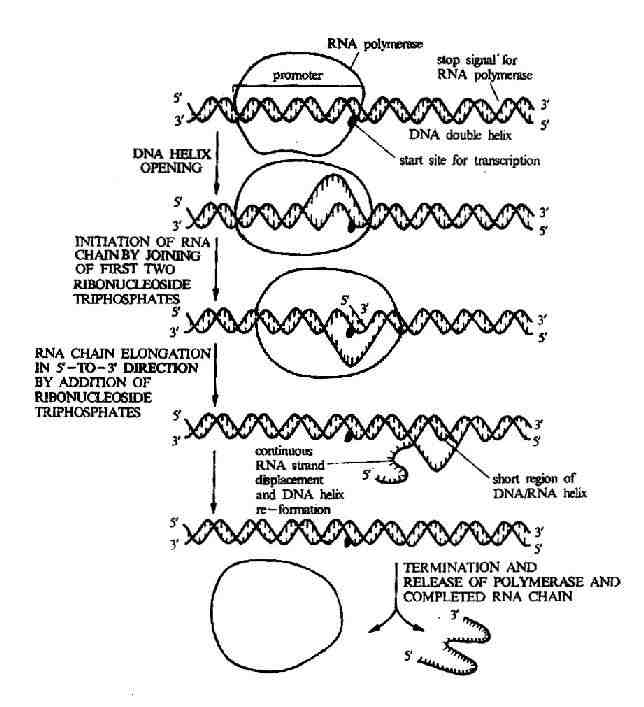
圖17-3 轉錄的主要過程
(一)識別
轉錄是從DNA分子的特定部位開始的,這個部位也是RNA聚合酶全酶結合的部信這就是啟動子。為什麼RNA聚合酶能夠僅在啟動子處結合呢?顯然啟動子處的核苷酸序列具有特殊性,為了方便,人們將在DNA上開始轉錄的第一個鹼基定為+1,沿轉錄方向順流而下的核苷酸序列均用正值表示;逆流而上的核苷酸序列均用負值表示。
對原核行物的100多個啟動子的序列進行了比較後發現;在RNA轉錄起始點上游大約-10bp和-35bp處有兩個保守的序列,在-10bp附近,有一組5』-TATAATpu的序列,這是Pribnow首先發現的稱為Pribnow框,RNA聚合酶就結合在互部位上。-35bp附近,有一組5』-TTGACG-的序列;已被證實與轉錄起始的辨認有關,是RNA聚合酶中的δ亞基識別並結合的位置。-35序列的重要性還在於在很大程度上決定了啟動子的強度。
由於RNA聚合酶分子很大,大約能覆蓋70bp的DNA序列,因此酶分子上的一個適合部位就能佔據從-35到-10序列區域(圖17-4)。
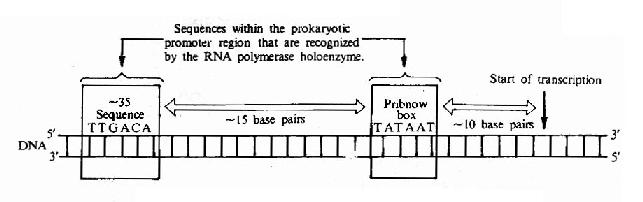
圖17-4 Structure of the prokaryotic promoter region.
真核生物的啟動子有其特殊性,真核生物有三種RNA聚合酶,每一咱都有自己的啟動子類型。以RNA聚合酶Ⅱ的啟動子結構為例,人們比較了上百個真核生物RNA聚合酶Ⅱ的啟動子核苷酸序列伯發現;在-25區有TATA框,又稱為Hogness框或Goldberg-Hogness框。其一致序列為T28A97A93A85A63T37A83A50T37,基本上都由A,T鹼基所組成,離體轉錄實驗表明,TATA框決定了轉錄起點的選擇,天然缺少TATA框的基本可以從一個以上的位點開始轉錄。在-75區有CAAT框,其一致的序列為GGTCAATCT。有實驗表明CAAT框與轉錄起始頻率有關,例如缺失GG,兔子的β珠蛋白基因轉錄效率只有原來的12%(圖17-5)。
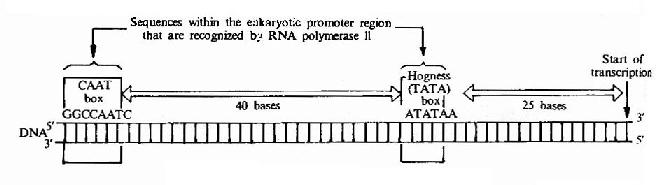
圖17-5 Eukaryotic gene promoter sequences
除啟動子外,真核生物轉錄起始點上游處還有一個稱為增強子的序列,它能極大地增強啟動子的活性,它的位置往往不固定,可存在於啟動子上游或下游,對啟動子來說它們正向排列和反向排列均有效,對異源的基因也起到增強作用,但許多實驗證實它仍可能具有組織特異性,例如免疫球蛋白基因的增強子只有在B淋巴細胞內活性最高,胰島素基因和胰凝乳蛋白酶基因的增喲子也都有很高的組織的特異性。
(二)轉錄起始和延伸
在原核生物中,當RNA聚合酶的δ亞基發現其識別位點時,全酶就與啟動子的-35區序列結合形成一個封閉的啟動子複合物。由於全酶分子較大,其另一端可在到-10區的序列,在某種作用下,整個酶分子向-10序列轉移並與之牢固結合,在此處發生局部DNA12-17r 的解鏈形成全酶和啟動子的開放性複合物。在開放性啟動子複合物中起始位點和延長位點被相應的核苷酸前體充滿,在RNA聚合酶β亞基催化下形成RNA的第一個磷酸二酸鍵。RNA合成的第一個核苷酸總有GTP或ATP,以GTP常見,此時δ因子從全酶解離下來,靠核心酶在DNA鏈上向下游滑動,而脫落的δ因子與另一個核心酶結合成全酶反覆利用。
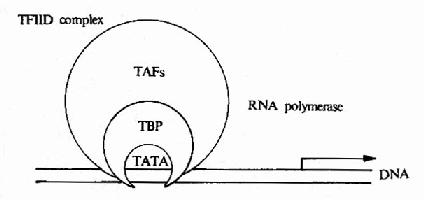
圖17-6 轉當起始複合物的模式
真核生物轉錄起始十分複雜,往往需要多種蛋白因子的協助,已經知道,在所有的細胞中有一類叫做轉錄因子的蛋白質分子,它們與RNA聚合酶Ⅱ形成轉錄起始複合物,共同參與轉錄起始的過程。
真核生物基因中,有專門為蛋白質編碼的基因,這些基因由RNA聚合酶Ⅱ負責進行轉錄起始關鍵性作用。根據這些轉錄因子的作用特點可大致分為二類;第一類為普遍轉錄因子它們與RNA聚合酶Ⅱ共同組成轉錄起始複合物,轉錄才能在正確的位置上開始。普遍轉錄因子是由多種蛋白質分子組成的,其中包括特異結合在TATA盒上的蛋白質,叫做TATA盒結合蛋白,還有至成一組複合物叫做轉錄因子ⅡD。TFⅡD再與RNA聚合酶Ⅱ結合完成轉錄起始複合物的形成。(圖17-6)
除TFⅡD以外,在細胞核提取物中還發現TFⅡA,TFⅡF,TFⅡE,TFⅡH等,它們在轉錄起始複合物組裝的不同階段起作用,像TFⅡH就有旋轉酶活性,它可利用ATP分解產生的能量,介導起始點雙螺旋的打開,使RNA聚合酶得以發揮作用。從中也不難看出真核細胞中基因轉錄的起始是基因的表達調控的關鍵,這麼多蛋白質分子之間相互作用,以及這些蛋白質分子DNA調控無件相結合,構成控制基因轉錄開始的複雜體系。
第二類轉錄因子為組織細胞特異性轉錄因子或者叫可誘導性轉錄因子,這此TF是在特異的組織細胞或是受到一些類固醇激素,生長因子或其它刺激後,開始表達某些特異蛋白質分子時,才需要的一類轉錄因子。
例如:激活劑蛋白-1就是一類可誘導的轉錄因子,它是由多蛋白質成份組成的複合物,可發由fos基因和jun基因家庭的蛋白質產物組成。當某些生長因子細胞因子和某些化學物質在細胞外刺激這些細胞時,使細胞內JUN蛋白和FOS蛋白髮生磷酸化,特異地結合到c-jun基因和c-fos基因的啟動子部位,使這些基因轉錄並翻譯出相應的c-JUN蛋白和c-FOS蛋白,這些蛋白質就可組成二聚體的AP-1,AP-1就會結合到細胞核中靶基因的調控部位,促進或激活靶基因的轉錄活性,產生出由於細胞外刺激因素作用下,這些細胞做出的特異反應一表達出的特異蛋白須分子。有關詳細內容見信號轉錄一章。
RNA鏈的延長靠核心酶的催化,在起始複合物上第一個GTP的核糖3』-OH上與DNA模板能配對的第二個磷酸核苷起反應形成磷酸二酯鍵。聚合進去的核苷酸又有核糖3』-OH游離,這樣就可按模板DNA的指引,一個接一個地延長下去。因此RNA鏈的合成方面也是5』--3。由於DNA鏈與合成的RNA鏈具有反平行關係,所以RNA聚合酶是沿著DNA鏈3』--5』方向移動。整個轉錄過程是由同一個RNA聚合酶來完成的一個連續下斷的反應,轉錄本RNA生成後,暫時與DNA模板鏈形成DNA.RNA雜交體,長度約為12個鹼基對,形成一個轉錄泡(圖17-7)。轉錄速度大允是每秒鐘30-50個核苷酸,但並不是以恆定速度進行的。在電子顯微鏡下觀察轉錄現象,可以看到同一DNA模板上,有長短不一的新合成的RNA鏈散開成羽毛狀圖形,這說明在同一DNA基因上可以有很鑫的RNA聚合酶在同時催化轉錄,生成相應的RNA鏈。而且較長的RNA鏈上已看到核糖體附著,形成多聚核糖體。說明某些情況下,轉錄過程未完全終止,即已開始進行翻譯。
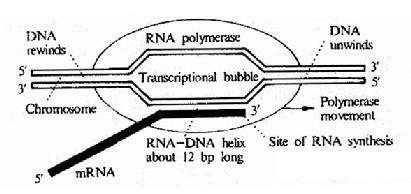
圖17-7 Diagrammatic representation of DNA transcription by E.coli RNApolymerase.The polymerase unwinds a stretch of DNA about 17base pairs in lengthforming a transcriptional bubble thatprogresses along thd DNA.The DNA has tounwind ahead of the polymerase and rewind behind it .The newly formed RNA formsa RNA-DNA double helix about 12 bae pairs long.
轉錄的延長階段,原核生物與真核生物之間沒有太大差別。
(三)轉錄的終止(Termination)
轉錄是在DNA模板某一位置上停止的,人們比較了若干原核生物RNA轉錄終止位點附近的DNA序列,發現DNA模板上的轉錄終止信號有兩種情況,一類是不依賴於蛋白質因子而實現的終止作用,另一類是依賴蛋白質輔因子才能實現終止作用,這種蛋白質輔因子稱為釋放因子,通常又稱ρ因子。兩類終止信號有共同的序列特徵,在轉錄終止之前有一段迴文結構,迴文序列是一段方向相反,鹼基互補的序列,在這段互補序列之間由幾個鹼基隔開,不依賴ρ因子的終止序列中富含G.C鹼基對,其下游6-8個A;而依賴ρ因子的終止序列中G.C鹼基對含量較少,其少游也沒有因固定的特徵,其轉錄生成的RNA可形成二級結構即柄一嚕噗結構,又稱髮夾結構,這樣的二級結構可能與RNA聚合酶某種特定的空間結構相嵌合,阻礙了RNA聚合酶進一步發揮作用(圖17-8)。除DNA模板本身的終止信號外,在入噬菌體中,發現一些蛋白質有協助RNA聚合酶跨越終止部位的作用,叫做抗轉錄終止蛋白,例如入噬菌體的N基因產物。
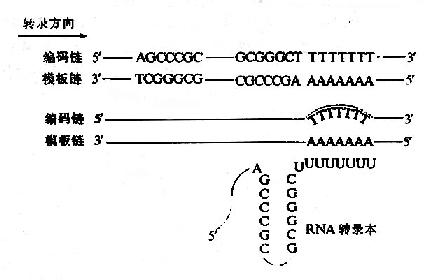
圖17-8 原核生物轉錄作用的終止信號
真核生物由於RNA轉錄後很快就進行了加工,因此很難確定原初轉錄物的3』末端。病毒SV40的終止位點經過研究發現,很像大腸桿菌的不依賴ρ因子的終止子,轉錄後的RNA可形成一個髮夾結構,3』末端帶有一連串的U。爪蟾5sRNA的3』末端有4個U,它們前後的序列為富含G.C的序列,這是所有真核生物RNA聚合酶Ⅲ轉錄的終止信號。這種序列特徵高度保守,從酵母到人都很相似,任何改變這種序列特徵的突變都將導致轉錄終止位置的改變。
| 關於「生物化學與分子生物學/RNA轉錄作用」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |