生理學/各段腎小管和集合管的轉運功能
| 醫學電子書 >> 《生理學》 >> 尿的生成和排出 >> 腎小管與集合管的轉運功能 >> 各段腎小管和集合管的轉運功能 |
| 生理學 |
|
|
|
(一)近球小管
腎小球濾過流經近球小管後,濾過液中67%Na+、CI-、K+和水被重吸收,85%的HCO3也被重吸收,葡萄糖、胺基酸全部被重吸收;H+則分泌到腎小管中。近球小管重吸收的關鍵動力是基側膜上的Na+泵;許多溶質,包括水的重吸收都與Na+泵的活動有關。
1.Na+、CI-和水的重吸收在近球小管前半段,大部分Na+與葡萄糖,胺基酸同向轉運、與H+逆向轉運而被主動重吸收;面近球小管後半段,Na+和CI-主要通過細胞旁路而被被動重吸收。水隨NaCI等溶質重吸收而被重吸收,因此,該段小管注與血漿滲透壓相同,是等滲重吸收。
在近球小管前半段,由於Na+泵的作用,Na+被泵至細胞間隙,使細胞內Na+濃度低,細胞內帶負電位。因此,小管液中的Na+和葡萄糖與管腔膜上的同向轉運體結合後,Na+順電化學梯度通過管腔膜的同時,釋放的能量將葡萄糖同向轉運入細胞內。進入細胞內的Na+即被細胞基側膜上的Na+泵泵出至細胞間隙,這樣,一方面使細胞內Na+的濃度降低,小管液中的Na+-葡萄糖便可不斷轉運進入細胞內,細胞內的葡萄糖由易化擴散通過細胞基側膜離開細胞回到血液中;另一方面,使細胞間隙中的Na+濃度升高,滲透壓也升高,通過滲透作用,水隨之進入細胞間隙。由於細胞間隙在管腔膜側的緊密連接相對是密閉的,Na+和水進入後就使其中的靜水壓升高,這一壓力可促使Na+和水通過基膜進入相鄰的毛細血管而被重吸收,但也可能使部分Na+和水通過緊密連接回漏(back-leak)至小管腔內(圖8-10A)。
另一部分的Na+-H+交換而主動重吸收。小管液中的Na+和細胞內的H+與管腔膜上的交換體結合進行逆向轉運,使小管液中的Na順濃度梯度通過管腔膜進入細胞的同時,將細胞內的H+分泌到小管液中;進入細胞內的Na+隨即被基側膜上的Na+泵泵至細胞間隙而主動重吸收。分泌到小管液中的H+將有利於小管液中的HCO3的重吸收。
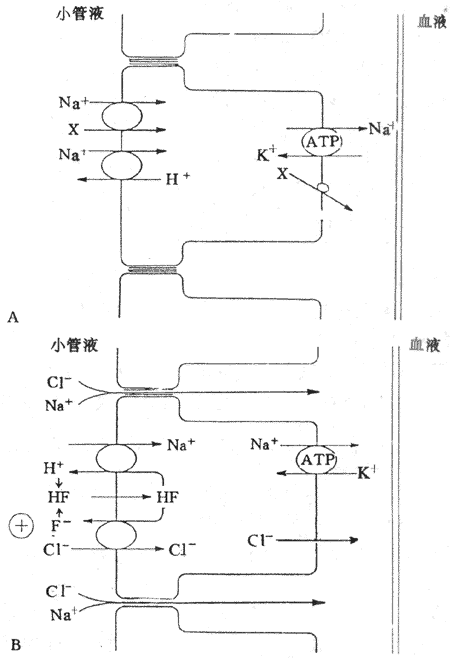
圖8-10 近球小管重吸收NaCI的示意圖
A:近球小管的前半段 X代表葡萄糖、胺基酸、磷酸鹽CI-
B:近球小管的後半段F-代表甲酸鹽 HF,甲酸
在近球小管後半段,NaCI是通過細胞旁路和跨上皮細胞兩條途徑而被重吸收的。小管液進入近球小管後半段時,絕大多數的葡萄糖、胺基酸已被重吸收。由於HCO3重吸收速率明顯大於CI-重吸收,CI-留在小管液中,造成近球小管後半段的CI-濃度比管周組織間液高20%-40%。因此,CI-順濃度梯度經細胞旁路(即通過緊密連接進入細胞間隙)而重吸收回血。由於CI-被動重吸收是生電性的,使小管液中正離子相對較多,造成管內外電位差,管腔內帶正電,管外帶負電,在這種電位差作用下,Na+順電位差通過細胞旁路而被動重吸收。CI-通過細胞旁路重吸收是順濃度梯度進行的,而Na+通過細胞旁路重吸收是順電位梯度進行的,因此,NaCI是重吸收都是被動的(圖8-10B)。
NaCI跨上皮細胞重吸收與H+和甲酸鹽(formate,F-)再循環有關,要通過管腔膜上相互耦聯的Na+-H+交換和CI—甲酸根交換。在正常腎小管液中含有低濃度甲酸根,通過Na+、H+交換,Na+進入細胞,H+分泌到小管液中,並與小管液中的甲酸根結合形成甲酸(formic acid,HF)。甲酸是脂溶性的,可迅速通過管腔膜進入細胞,在細胞內分解為H+和甲酸根。甲酸根和小管液中的CI-進行逆向轉運,結果,CI-進入細胞內並通過基側膜而被重吸收,而甲酸根則人細胞內進入小管液。細胞內的H+則與小管液中的Na+進行逆向交換,Na+進入細胞,並被Na+泵泵至細胞間隙,然後進入管周毛細血管而被重吸收;H+分泌至小管液,再與小管液中的甲酸根據結合,形成甲酸再進入細胞。因此,H+和甲酸根可再循環使用,CI-和Na+則被重吸收回血。
水的重吸收是被動的,是靠滲透作用而進行的。水重吸收的滲透梯度存在於上皮細胞和細胞間隙之間。這是由於Na+順電化學梯度通過管腔膜進入細胞後,細胞內的Na+被基側膜上的Na+泵泵至細胞間隙,使細胞間隙滲透壓升高。在滲透作用下,水便不斷從小管液進入上皮細胞,並從細胞不斷進入細胞間隙,造成細胞間隙靜水壓升高;加上管周毛細血管內靜水壓較低,膠體滲透壓較高,水便通過周圍組織間隙進入毛細血管而被重吸收。
2.HCO3重吸收與H+的分泌 HCO3的重吸收與小管上皮細胞管腔膜上的Na+-H+交換有密切關係。HCO3在血漿中鈉鹽(NaHCO3)的形式存在,濾過中的NaHCO3濾入囊腔進入腎小管後可解離成Na+和HCO3。通過Na+-H+交換,H+由細胞內分泌到小管液中,Na+進入細胞內,並與細胞內的HCO3一起被轉運回血(圖8-11)。由於小管液中的HCO3不易通過管腔膜,它與分泌的H+結合生成H2CO2,在碳酸酐酶作用下,H2CO2迅速分解為CO2和水。CO2是高度脂溶性物質,能迅速通過管腔膜進入細胞內,在碳酸酐酶作用下,進入細胞內的CO2與H2O結合生成H2CO3。H2CO3又解離成H+和HCO3。H+通過Na+-H+交換從細胞分泌到小管液中,HCO3則與Na+一起轉運回血。因此,腎小管重吸收HCO3是以CO2的形式,而不是直接以HCO3的形式進行的。如果濾過的HCO3超過了分泌的H+,HCO3就不能全部(以CO2形式)被重吸收。由於它不易透過管腔膜,所以餘下的便隨尿排出體外。可見腎小管上皮細胞分泌1H+就可使1HCO3和1Na+重吸收回血,這在體內的酸鹼平衡調節中起到重要作用。乙醯唑胺可抑制碳酸酐酶的活性,因此,用乙醯唑受後,Na+-H+交換就會減少,因而NaHCO3、NaCI和水的排出增加,可引起利尿。由於近球小管的Na+-H+交換,小管液中的HCO3與H+結合併生成CO2,CO2透過管腔膜的速度明顯高於CI-的速度。因此,HCO3的重吸收率明顯大於CI-的重吸收率。
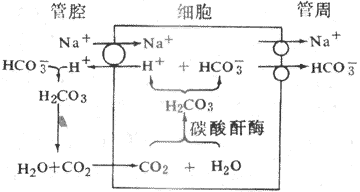
圖8-11 腎小管上皮細胞生成和分泌H+示意圖
3.K+的重吸收微穿刺實驗表明,腎小球濾過的K+,67%左右在近球小管重吸收回血,而尿中的K+主要是由遠曲小管和集合管分泌的。有人認為,近球小管對K+的重吸收是一個主動轉運過程。小管液中鉀濃度為4mmol/L,大大低於細胞內K+濃度(150mmol/L)。因此在管腔膜處K+重吸收是逆濃度梯度進行的。管腔膜K+主動重吸收的機制尚不清楚。
4.葡萄糖重吸收腎小球濾過液中的葡萄糖濃度與血糖濃度相同,但尿中幾乎不含葡萄糖,這說明葡萄糖全部被重吸收因血。微穿刺實驗表明,重吸收葡萄糖的部位僅限於近球小管,尤其是在近球小管前半段,其他各段腎小管都沒有重吸收葡萄糖的能力。因此,如果在近球小管以後的小管液中仍含有葡萄糖,則尿中將出現葡萄糖。
葡萄糖是不帶電荷的物質,它的逆濃度梯度重吸收的,是由Na+繼發性主動同向轉運而被重吸收的。在兔腎近球小管微灌流實驗中觀察到,如果灌流液中去掉葡萄糖等有機溶質,則Na+的重吸收率降低;如果灌流液中的Na+全部去掉,則葡萄糖有機溶質的重吸收將完全停止,說明葡萄糖的重吸收與Na+同向轉運密切相關。葡萄糖和Na+分別與管腔膜上的同向轉運體蛋白的結合位點相結合而進行同向轉運(見前述Na+重吸收)。
近球小管對葡萄糖的重吸收有一定限度。當血液中葡萄糖濃度超過160-180mg/100ml時,有一部分腎小管對葡萄糖的吸收已達到極限,尿中開始出現葡萄糖,此時的血糖濃度稱為腎糖閾。血糖濃度再繼續升高,尿中葡萄糖含量也將隨之不斷增加;當血糖濃度超過300mg/100ml後,全部腎小管對葡萄糖的吸收均已達到極限,此值即為葡萄糖吸收極限量。此時,尿葡萄糖排出率則隨血糖濃度升高而平行增加。人腎的葡萄糖吸收極限量,在體表面積為1.73m2的個體,男性為375mg/min,女性為300mg/min。腎之所以有葡萄糖吸收極限量,可能是由於同向轉運體的數目有限的緣故,當所有同向轉運體的結合位點都被結合而達飽和時,葡萄糖轉運量就無從再增加了。
5.其他物質的重吸收和分泌小管注保的胺基酸的重吸收與葡萄糖的重吸收機制相同,也與Na+同向轉運(圖8-9)。但是,轉運葡萄糖的和轉運胺基酸的同向轉運體可能不同,也就是說同向轉運體具有特異性。此外,HPO4-2、SO4-2的重吸收也也Na+同向轉運而進行。正常時進入濾液中的微量蛋白質則通過腎小管上皮細胞吞飲作用而被重吸收。
體內代謝產物和進入體仙的某些物質如青黴素、酚紅,大部分的利尿藥等,由於與血漿中蛋白結合而不能通過腎小球濾過,它們均在近球小管被主動分泌到小管液中而排出體外。
(二)髓袢
近球小管液流經髓袢過程中,約20%的Na+、CI-、和K+等物質被進一步重吸收。髓袢升支粗段的NaCI重吸收在尿液稀釋和濃縮機制中具有重要意義。髓袢升支粗段CI-是逆電化學梯度被上皮細胞重吸收的。微穿刺實驗證明,兔髓袢升支粗段管腔內為正電位(+10mV)。在微灌流實驗中,如果灌流液中不含K+,則管內的正電位基本消失,CI-重吸收率很低,這說明管腔內正電位與CI-的重吸收和小管液中的K+有密切關係。如果在髓袢升支粗段管周的浸溶液中加入選擇性Na+泵抑制劑哇巴因(ouabain)抑制Na+泵後,CI-的轉運也受阻,說明Na+泵是CI-重吸收的重要因素。據上述實驗,有人提出Na+:2CI-:K+同向轉運模式來解釋升支NaCI的繼發性主動重吸收。該模式認為:①髓袢升支粗段上皮細胞基側膜上的Na+泵,將Na+由細胞內示向組織間液,使細胞內的Na+下降,造成管腔內與細胞內Na+有明顯的濃度梯度;②Na+與管腔膜上同向轉運體結合,形成Na+:2CI-:K+同向轉運體複合物,Na+順電化學梯度將2CI-和K+一起同向轉運至細胞;③進入細胞內的Na+、2CI-和K+的去向各不相同:Na+由Na+泵泵至組織間液,2CI-由於濃度梯度經管周膜上CI-通道進入組織間液,而K+則順濃度梯度經管腔膜而返回管腔內,再與同向轉運體結合,繼續參與Na+:2CI-K+的同向轉運,循環使用;④由於2CI-進入組織間液,K+返回管腔內,這就導致管腔內出現正電位;⑤由於管腔內正電位,使管腔液中的Na+等正離子順電位差從細胞旁路進入組織間液,這是不耗能的Na+被動重吸收。從這個模式說明,通過Na+泵的活動,繼發性主動重吸收了2CI-,同時伴有2Na+的重吸收,其中1Na+是主動重吸收,另1Na+通過細胞旁路而被動重吸收,這樣為Na+的重吸收節約了50%能量消耗(圖8-12)。髓袢升支粗段對水的通透性很低,水不被重吸收而留在小管內。由於NaCI被上皮細胞重吸收至組織間液,因此造成小管液低滲,組織間液高滲。這種水和鹽重吸收的分離,有利於尿液的濃縮和稀釋。Na+:2CI-:K+同向轉運對速尿,利尿酸等利尿劑很敏感。它們與同向轉運體結合後,可抑制其轉運功能,管腔內正電位消失,NaCI的重吸收受抑制,從而干擾尿的濃縮機制,導致利尿。
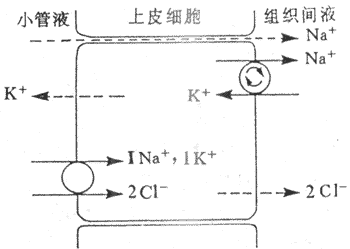
圖8-12 髓袢升支粗段繼發性主動吸收CI-的示意圖
(三)遠球小管和集合管
在遠曲小管和集合管,重吸收大約12%濾過的Na+和CI-,分泌不同量的K+和H+,重吸收不同量的水。水、NaCI的重吸收以及K+和H+的分泌可根據機體貼的水、鹽平衡狀況來進行調節。如相機缺水或缺鹽時,遠曲小管和集合管可增加水、鹽的重吸收;當機體水、鹽過剩時,則水、鹽重吸收明顯減少,水和鹽從尿排出增加。因此,遠曲小管和集合管對水和鹽的轉運是可被調節的。水的重吸收主要受抗利尿激素調節,而Na+和K+的轉運主要受醛固酮調節。
遠曲小管和集合管上皮細胞間隙的緊密連接對小離子如Na+、K+和CI-等的通透性低,這些離子不易通過緊密連接回漏至小管腔內,因此,所能建立起來的管內外離子濃度梯度和電位梯度大。在遠曲小管初段,對水的通透性很低,但仍主動重吸收NaCI,繼續產生低滲小管液。Na+在遠曲小和集合管的重吸收是逆較大的電化學梯度進行的,是主動重吸收過程。這可能與遠曲小管的Na+泵在腎單位中活性最高有關。有人認為在遠曲小管初段的小管液中,Na+是通過Na+-CI-同向轉運進入細胞的,然後由Na+泵將Na+泵出細胞而主動重吸收回血(圖8-13A)。NaCI同向轉運體可被噻嗪類利尿藥所抑制。
遠曲小管後段和集合管含有兩類細胞,即主細胞和閏細胞。主細胞重吸收Na+和水,分泌K+,閏細胞則主要分泌H+,主細胞重吸收Na+主要通過管腔膜上的Na+通道。管腔內的Na+順電化學梯度通過管膜上的Na+通道進入細胞,然後,由Na+泵泵至細胞間液而被重吸收(圖8-13B)。
K+的分泌 尿中K+的排泄量視K+的攝入量而定,高鉀飲食可排出大量的鉀,低鉀飲食則尿中排鉀量少,使機體的鉀攝入量與排出量保持平衡,維持機體K+濃度的相對恆定。
K+分泌的動力包括:①在遠曲小管和集合管的小管液中,Na+通過主細胞的管腔膜上的Na+通道進入細胞,然後,由基側膜上的Na+泵將細胞內的Na+泵至細胞間隙而被重吸收,因而是生電性的,使管腔內帶負電位(-10?40mV)。這種電位梯度也成為K+從細胞分泌至管腔的動力;②在遠曲小管後段和集合管的主細胞內的K+濃度明顯高於小管液中的K+濃度,K+便順濃度梯度從細胞內通過管腔膜上的K+通道進入小管液;③Na+進入主細胞後,可刺激基側膜上的Na+,使更多的K+從細胞外液中泵入細胞內,提高細胞內K+濃度,增加細胞內和小管液之間的K+濃度梯度,從而促進K+分泌,因此,K+的分泌與Na+的重吸收有密切關係。(圖8-13B)。
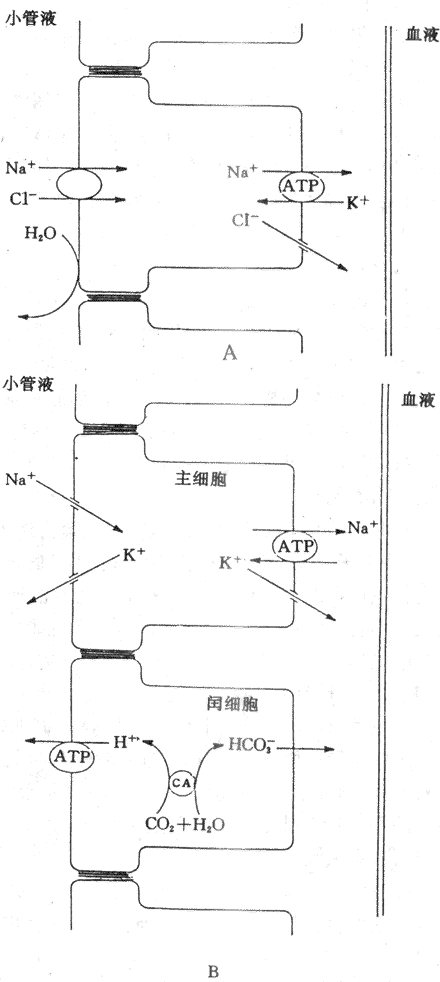
圖8-13 遠球小管和集合管重吸收NaCI、分泌K+和H+的示意圖
A:遠曲小管初段 B:遠曲小管後段和集合管
H+的分泌除了近球小管細胞通過Na+-H+交換分泌H+,促進NaHCO3重吸收外,遠曲小管和集合管的閏細胞也可分泌H+。H+的分泌是一個逆電化學梯度進行的主動轉運過程。有人認為管腔膜上有H+泵,能將細胞內的H+歷史意義入小管腔內。細胞內的CO2和H2O在碳酸酐酶催化作用下生成的H+和HCO3,H+由H+泵泵至小管液,HCO3則通過基側膜回到血液中,因而H+分泌和HCO3的重吸收與酸鹼平衡的調節有關(圖8-13B)。閏細胞分泌的H+與HPO4-2結合形成H2PO4,這是可滴定酸;分泌的H+可與上皮細胞分泌的NH3結合,形成NH4+。可滴定酸和NH4+都不易透過管腔膜進入細胞而留在小管液中。因此,它們是尿液酸鹼度的決定因素。
NH3的分泌遠曲小管和集合管的上皮細胞在代謝過程中不斷地生成NH3,這些NH3主要由谷氨醯胺脫氨而來。NH3具有脂溶性,能通過細胞膜向小管液周圍組織間液和小管液自由擴散,擴散量取決於兩種液體的pH值。小管液的pH較低(H+濃度較高),所以NH3較易向小管液中擴散。分泌的NH3能與小管液中的H+結合併生成NH4-,小管液中NH3濃度因而下降,於是管腔膜兩側形成了NH3濃度梯度,此濃度梯度又加速了NH3向小管液中擴散。由此可見,NH3的分泌與H+的分泌密切相關;H+分泌增加促使NH3分泌增多。NH3與H+結合併生成NH4-後,可進一步與小管液中的強酸鹽(如NaCI等)的負離子結合,生成酸性銨鹽(NH4CI等)並隨尿排出。強酸鹽的正離子(如Na+)則與H+交換而進入腎小管細胞,然後和細胞內HCO3-一起被轉運回血。所以,腎小管細胞分泌NH3,不僅由於銨鹽形成而促進了排H+,而且也促進了NaHCO3的重吸收。
| 關於「生理學/各段腎小管和集合管的轉運功能」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |