生物化學與分子生物學/細胞癌基因的激活
| 醫學電子書 >> 《生物化學與分子生物學》 >> 癌基因與抑癌基因 >> 細胞癌基因的激活 |
| 生物化學與分子生物學 |
|
|
|
細胞癌基因的激活是指原本不致癌c-onc在特定的情況下轉變成致癌性的,大體上有以下幾種激活方式。
1、插入激活 例如逆轉錄病毒MoSV感染鼠類成纖維細胞後,病毒基因組的LTR整合到細胞癌基因c-mos鄰近處,使c-mos處於LTR的強啟動子和增強子作用之下而被激活,導致成纖維細胞轉化為肉瘤細胞,又如禽類白細胞增生病毒ALV的E成分整合到雞細胞基因組c-myc附近。可使c-myc激活。因此在基因治療中使用逆轉錄病毒載體時必需考慮細胞癌基因的插入激活問題。
2、突變激活 典型的是各種ras基因的激活,參看錶22-2。
| ras基因 | 胺基酸的位置 | |||
| 12 | 13 | 59 | 61 | |
| H-ras-1來源的 正常人c-H-ras-1 人EJ膀胱癌症 人HS242乳癌 鼠Harvey病毒v-H-ras |
Gly GGC Val GTC Arg CGC |
Gly | Ala Thr |
Gln CAG Leu CTG |
| K-ras-2來源的 正常人c-K-ras-2 人Caalu肺癌 人SW480結腸癌 鼠Kirsten病毒v-K-ras |
Gly Lys TGT Val GTT Ser |
Gly | Ala Thr |
Gln |
| N-ras來源的 正常人N-ras 人神經母細胞瘤 人早幼粒細胞白血病細胞系 人纖維肉瘤 人AML33 |
Gly | Gly Asp |
Ala | Gln Lys Lys Lys |
ras基因的表達產物Ras是一種小分子G蛋白,在信號轉導中起重要作用,正常Ras的作用因其自身的GTP酶活性而受到嚴格控制,而突變了的Rad其GTP酶活性下降或喪失,失去了原有控制,致使增殖信號持續作用,細胞發生惡性轉化,如圖22-3所示。
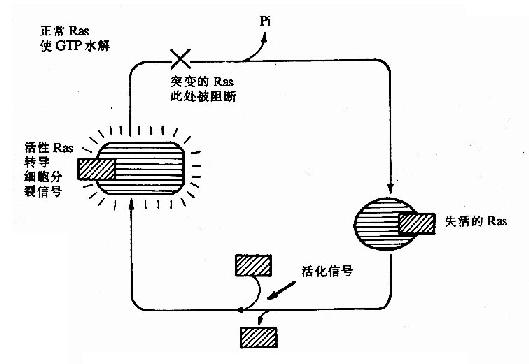
圖22-3 Ras與GTP/GDP的相互作用及突變的RasR的作用
表22-3 人類腫瘤細胞中擴增的細胞癌基因
| c-onc | 腫瘤 | 擴增倍數 | DM/HSR* |
| c-myc | 早幼粒白血病細胞系HL60 | 20× | + |
| 小細胞肺癌細胞系 | 5-30× | ? | |
| N-myc | 原發神經母細胞瘤Ⅲ-Ⅳ級及神經母細胞瘤細胞系 | 5-1000× | + |
| 視網膜母細胞瘤 | 10-200× | + | |
| 小細胞肺癌 | 50× | + | |
| L-myc | 小細胞肺癌 | 10-20× | ? |
| c-myb | 急粒AML | 5-10× | ? |
| 結腸癌細胞系 | 10× | ? | |
| c-erbB | 類表皮癌細胞系,原髮膠質瘤 | 30× | ? |
| c-K-ras | 原發肺癌,結腸癌,膀胱癌,直腸癌 | 4-20× | ? |
| N-ras | 乳癌細胞系 | 5-10× | ? |
典型的如伯基特淋巴瘤細胞的染色體易位t(8:14),致使c-myc激活,參看錶22-4和圖22-4。
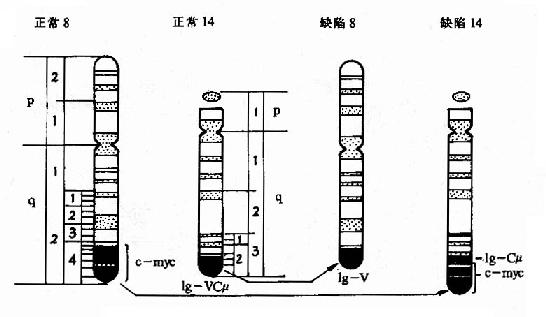
圖22-4 Burkitt淋巴瘤常見的染色體易位t(8:14)
表22-4 染色體異常與癌基因重排
| 癌基因 | 染色體定位 | 異常 | 人類腫瘤 |
| c-myc | 8q24 | t(8:14),t(8:22) | Burkitt淋巴瘤 |
| t(2:8) | |||
| bcl-1 | 11q13 | t(11:14) | B細胞淋巴瘤 |
| bcl-2 | 18q21 | t(14:18) | |
| tcl-2 | 11q13 | t(11:14) | T細胞淋巴瘤 |
| c-abI | 9q34 | t(9:22) | 慢粒CML |
| bcr | 22q11 | ph | |
| c-mos | 8q22 | t(8:21) | 急粒AML |
| c-myb | 6q22-24 | t(6:14) | 卵巢癌 |
| c-sis | 22q12 | t(11:22) | Erwing網瘤 |
| blym | 1q32-ter | 缺失,HSR | 神經纖維瘤 |
| c-K-ras | 6q21 | 斷裂 | ANLL |
| 6q三體性 | 視網膜母細胞癌 | ||
| c-erbA | 17q21 | 斷裂 | ANLL |
已知B淋巴細胞中免疫球蛋白重鏈基因表達十分活躍,其啟動子為強啟動子,且在CH-VH之間還有增強子區,c-myc易位後與IG重鏈基因的調控區為鄰,因而被激活。正常情況下,位於c-myc5』端的兩個啟動子受到c-myc產物的反饋抑制,由此重排時5』端序列有丟失,結果擺脫了抑制而表達增強。
不同的癌基因有不同的激活方式,一種癌基因也可有幾種激活方式。例如c-myc的激活就有基因擴增和基因重排兩種方式,很少見c-myc的突變;而ras的激活方式則主要是突變,1985年Slamon檢測了20種54例人類腫瘤中的15種癌基因,發現所有腫瘤都不止一種癌基因發生改變。細胞轉化實驗證明,各種癌基因之間存在協同作用。例如,單獨v-myc或EJ-ras都不能使大鼠胚胎成纖維細胞轉化,但是若將二者共轉染PEF,8天後80%的細胞發生變化,那麼為什麼單獨EJ-ras又可使Rat-1細胞轉化呢?原因是該細胞並非正常,而是已經永生化了的細胞,如果先用化學誘癌物或射線使正常大鼠原代成纖維細胞永生化,然後再用EJ-ras轉染,則可使之轉化,因此Weingerg按轉染細胞表型的變化將癌基因分為兩個類,一類是核內作用的能使細胞永生化的癌基因,例如myc,fos等,另一類是引起細胞惡性表型變化的定位於質膜和胞漿的癌基因,例如ras、erbB、src等。事實表明腫瘤的發生是多步驟,多因素的,不同的癌基因作用於腫瘤發生的不同階段。
不僅癌基因之間有協同作用,癌基因與抑癌基因之間也存在協同作用。
| 關於「生物化學與分子生物學/細胞癌基因的激活」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |