生物化學與分子生物學/癌基因的發現
| 醫學電子書 >> 《生物化學與分子生物學》 >> 癌基因與抑癌基因 >> 癌基因的發現 |
| 生物化學與分子生物學 |
|
|
|
現已知道在腫瘤發生中,作為環境因素的病毒、化學致癌物和射線,它們作用於機體內的靶分子都是DNA,在研究腫瘤病毒如何使宿主細胞轉化和研究腫瘤DNA能否使培養的經兩條實驗途徑中,殊途同歸,發現了癌基因,早在本世紀初,Rockefeller研究所的Rous醫生將雞肉瘤組織勻漿後的無細胞濾液皮下注射於正常雞,發現可以引起腫瘤,可惜當時對病毒還缺乏認識,直到五十年代才重新發現原來致瘤的因素是病毒,並以Rous醫生的名字命名為羅氏肉瘤病毒(Rous Sarcoma Virus ,RSV)。1975年,Bishop從RSV中分離到第一個病毒癌基因src,該基因編碼分子量為60kDa的磷蛋白質,以pp60src表示。
1976年Stehelin以實驗證明正常雞成纖維細胞基因組中存在有與病毒癌基因src的同源序列。此後陸續發現許多禽類和鼠類病毒部基因也有類似情況,即宿主細胞基因組中含有病毒癌基因的同源序列,稱之為細胞癌基因(c-oncogene,c-onc)。
那麼,v-onc與c-onc的關係如何?這可從二者結構的比較和逆轉錄病毒感染宿主後的生活史或複製周期兩方面加以分析。
首先,從結構上看c-onc是間斷的,這是真核基因的特點,即有內含子因而基因的跨度較大。然而v-onc卻是連續的,沒有內含子,所以基因跨度較小,以v-onc和c-src為例如圖22-1。再從逆轉錄病毒感染宿主細胞後的複製周期(圖22-2)分析。
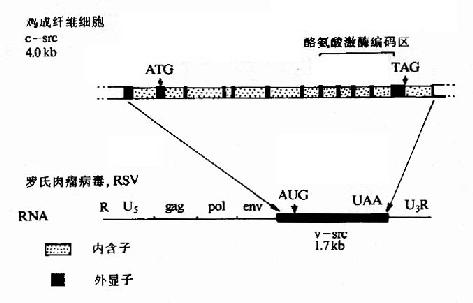
圖22-1 c-src和v-src結構的比較
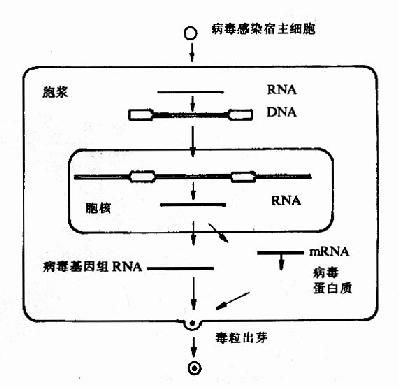
圖22-2 逆轉錄病毒正常複製周期主要步驟
不難看出,v-onc原本不是病毒的基因,而是動物細胞正常基因的一個複本。當病毒在宿主細胞內複製時,由於DNA重組而將宿主細胞基因中帶有v-onc的序列重組到病毒的基因組內。所以說c-onc是v-onc原型,又稱為原癌基因(proto-oncogene)。
病毒癌基因對病毒本身無關緊要,卻可使宿主細胞轉化,引起腫瘤,而細胞癌基因對細胞的生長、分化和功能活動卻是至關緊要的。正常的細胞癌基因並不致癌,只是當它們異常表達或其表達產物異常時才會導致細胞的惡性轉化,迄今發現的細胞癌基因都是一些有十分重要的功能「看家基因」,而且是高度保守的,例如人與小鼠的K-ras基因產物K-Ras的胺基酸序列相差僅為1%,人與大鼠的H-ras基因產物H-Ras的胺基酸序列完全相同。
逆轉錄病毒屬於RNA腫瘤病毒,在DNA腫瘤病毒的基因組也存在著能使宿主細胞轉化的基因,例如腺病毒的E1A、E1B基因,多瘤病毒的大T、中T基因。人乳頭瘤病毒的E6、E7基因,以及DV40中的大T基因。它們為病毒複製所必需,同時又有使宿主細胞轉化的作用,故沿用原名,不另以癌基因命名。
細胞癌基因可按其表達產物的功能和定位分類如表22-1。
表22-1 細胞癌基因按其產物定位和功能分類
| 定位 | 功能 | 癌基因,產物 |
| 分泌蛋白 | 生長因子 | sis PDGFβ鏈 |
| 跨膜蛋白 | 受體型酪氨酸激酶 | erb B ,EGF受體 erb B2 ,EGF樣受體 (neu) fms ,CSF-1受體 |
| 膜結合蛋白 | G-蛋白 非受體型酪氨酸激酶 |
ras p21ras src pp60src |
| 胞漿可溶性蛋白 | 非受體型酪氨酸激酶 絲氨酸/蘇氨酸激酶 信號轉導連接蛋白 |
ab1 raf crk,SH2/SH3調節蛋白 vav ,SH2調節蛋白 |
| 胞核蛋白 | 轉錄因子 | myc myb fos jun erb A,T3受體 |
從上表可以看出,所有細胞癌基因均在信號轉導中扮演著重要角色。正因為如此,它們的異常或「變質」--表達失控或表達產物異常--對細胞的危害就很大,既然v-onc只是c-onc的一個複本,那麼為什麼c-onc本不致癌而v-onc又會致癌呢?實驗證明,v-onc的致癌或由於表達的失控,或由於基因的突變,導致產物的量的增多或質的改變。
已知從自然發生的人腫瘤組織提取的DNA可以轉化HIH/3T3細胞,儘管只有10%的人的腫瘤DNA具有轉化此種細胞的能力,但癌基因已在所有主要類型人腫瘤中檢出,最先是從T24/EJ膀胱癌細胞系檢查到的,屬於ras家族成員,以後又用核酸探針檢測出正常人的細胞基因組中有ras同源序列存在,與T24細胞中的ras不同,無轉化能力,二者差別僅僅在於一個點突變(第12位胺基酸密碼子的G突變為T)。
| 關於「生物化學與分子生物學/癌基因的發現」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |