醫學免疫學/T細胞主要表面分子
| 醫學電子書 >> 《醫學免疫學》 >> 免疫細胞(二):淋巴細胞系 >> T細胞 >> T細胞主要表面分子 |
| 醫學免疫學 |
|
|
|
T細胞是由一群功能不同的異質性淋巴細胞組成,由於它在胸腺內分化成熟故稱為T細胞。成熟T細胞由胸腺遷出,移居於周圍淋巴組織中淋巴節的副皮質區和脾白髓小動脈的周圍。不同功能成熟的T細胞均屬小淋巴細胞,在形態學上不能區分,但可借其細胞膜表面分子不同加以鑒別(表8-1)。
在T細胞發育不同階段以及成熟T細胞在靜止期和活化期,其細胞膜分子表達的種類和數量均不相同。這些分子為抗原性不同的糖蛋白。它們與T細胞對抗原的識別、細胞的活化、信息的傳遞、細胞的增殖和分化以及T細胞的菜單達相關。它們也與T細胞在周圍淋巴組織中的定位相關。
由於這些分子在T細胞表面相當穩定,故可視為T細胞的表面標誌,可以用以分離。鑒定不同功能的T細胞。這些分子的單株抗體對臨床相關疾病的診斷和治療也具有重要應用價值。
表8-1 T細胞主要表面分子
| 名稱 | 生化特性 | 配體 | 功 能 粘附 信號傳導 |
| TCR | αβ異二聚γδ異二聚體 | MHC-肽複合分子 | + + |
| CD3 | 五聚體 | - | - + |
| CD4 | 單體分子 | MHCⅡ類分子 | + + |
| CD8 | 雙體分子 | MHCⅠ類分子 | + + |
| CD28 | 同二聚體分子 | B7/BB1 | + + |
| CD2(LFA-2) | 單體分子 | CD58(LFA-3) | + + |
| CD11α/CD18 (LFA-1) |
αβ異二聚體分子 | CD54 (ICAM-1) (ICAM-2) |
+ + |
| CDw49/CD29 (VLA-4、5、6) |
αβ異二聚體分子 | VCAM-1 | + ? |
| CD44)(Pgp-1) | 單體分子 | ECM | + + |
| CD45 | 單體分子 | ? | ? + |
(一)T細胞抗原識別受體(TCR)
1.TCRαβ TCR是T細胞識別蛋白抗原的特異性受體,不同的T細胞克隆其抗原識別受體的分子結構也是不相同的。大多數成熟T細胞(約佔95%)的TCR分子是由α和β二條異二聚體肽鏈組成的TCRαβ分子。二條肽鏈都由膜外區、穿膜區及胞漿區組成。TCR屬於Ig超家族,膜外區可包括可變區(V區)及穩定區(C區)。
編碼人TCRα鏈和β鏈基因座分別定位於第14號和7號染色體。α鏈是由V、J、C基因段編碼的肽鏈。每個基因座又各有不同的等位基因,在T細胞發育分化早期與Ig基因一樣經歷基因重排、轉錄和轉譯成為肽鏈。TCR的特異性是由α鏈和β鏈的V-J及V-D-J基因片段決定的,故二條鏈基因重排後可形成千萬種不同特異性的TCR分子,故可識別環境中多種多樣的抗原。在通常情況下,異種蛋白抗原分子必須與細胞表面的自身MHC分子結合才能TCR識別。所以TCR只能識別細胞膜上的MHC分子與抗原分子,這是與B細胞識別原的主要不同特性。
2.TCRγδ另一種TCR是由γ和δ鏈組成的TCRγδ分子,它是由γ和δ基因編碼的分子。這種TCRγδ細胞多見一胸腺內早期T細胞(CD4-,CD8-,TCRγδ+),而在人周圍血成熟T細胞(CD3+,TCRγδ+)中所佔的比例甚少,約為1%~10%。在小鼠脾、表皮細胞和腸粘膜上皮細胞中亦可發現γδ+T細胞。對這種新發現的T細胞的生理功能尚不不清楚,但它們可能是具有原始受體的第一防線的防禦細胞,與清除表皮及上皮細胞內異物有關,它們可能是具有原始受體的第一防線的防禦細胞,與清除表皮及上皮細胞內異物有關。它們可識別高度保守的抗原,如結核桿菌、腸毒素和熱休克蛋白等抗原,在人和小鼠均表明它們可識別MHC或MHC樣分子。
(二)CD3分子
此分子可表達於所有成熟T細胞表面,它是由五條肽鏈非共價結合組成的複合分子,分別稱為γ、δ、ε、ζ和η鏈。五條肽鏈均由胞外區、穿膜區和胞漿區組成。γ、δ和ε為單體,ζ和η鏈其胞外區可由雙硫鍵連接組成為同二聚體ζζ(約佔90%)和異二聚體ζη分子(約佔10%)。
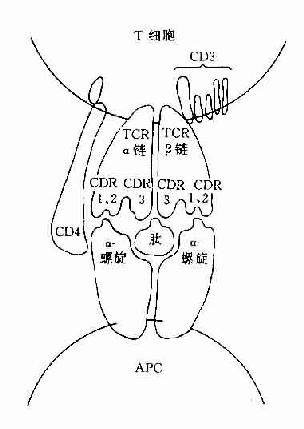
圖8-1 TCR識別普通抗原
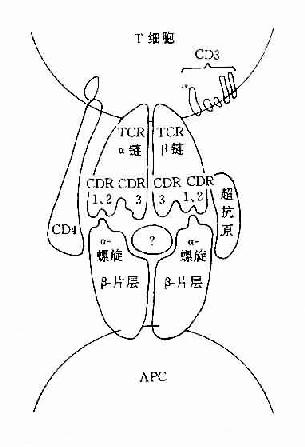
圖8-2 TCR識別超抗原
CD3分子可與TCR分子以非共價結合形成一個TCR-CD3複合受體分子,是T細胞識別抗原的主要識別單位。其中TCR是識別異種抗原和自身MHC分子多態性決定族的受體,而CD3分子交不參予抗原識別,它具有穩定TCR結構和傳遞活化信號的作用。
(三)CD4和CD8分子
這二種分子可同時表達於胸腺內早期胸腺細胞,稱為雙陽性胸腺細胞(CD4+、CD8+,DP)。而在成熟T細胞這二種分子是互相排斥的,只能表達一種分子,故可將成熟T細胞分為二類,即CD4+細胞和CD8+細胞。在外周淋巴組織中CD4+T約佔65%,CD8+T約佔35%。
這二種分子同屬於Ig超家族,都不具有多樣性。其分子結構都由胸外區、穿膜區及胸內區組成。CD4分子為55KD的單體,CD8分子為34KD多肽組成的雙體分子。
這二種分子與抗原識別無關,但可與帶有MHC分子的細胞結合,它們是細胞與細胞間相互作用的粘附分子。CD4分子是MHCⅡ類分子的受體,它可與MHCⅡ類分子的非多態區結合。CD8分子可與MHCⅠ類分子的非多態區結合。因此這二種分子具有增強TCR與抗原呈遞細胞或靶細胞的親和性,並有助於激活信號的傳遞。
(四)CD28分子
這種分子可表達於全部CD4+T細胞及50%CD8+細胞。它是80~90KD的由雙硫鍵連接的同源二聚體分子,屬Ig基因超家族。
近年的研究證明T細胞的活化需要雙信號,即由TCR-CD3複合分子可提供起始信號或第1信號,還必須有協同刺激信號(costimulatouy signal)或第2信號才能使T細胞活化。在T細胞膜上已發現有多種分子與協同刺激信號產生有關,如CD2、LFA-1、VLA-4及CD28分子等。稱這種分子為輔助分子或協同刺激受體分子。
其中以CD28分子最為重要,已證明它的配體分子存在於B細胞或其它抗原呈遞細胞上,命名為B7或BB1分子。它是50KD單體分子的穿膜蛋白,也屬Ig基因超家族。B7/BB1分子在靜止期B細胞、巨噬細胞或樹突狀細胞等表達弱,而活化型細胞表達增強。
(五)CD2分子
此分子亦稱為LFA-2、Len-5或羊紅細胞受體等名稱。為55KD單體分子,屬Ig基因超家族,亦為穿膜糖蛋白分子。可存在於成熟T細胞及胸腺細胞,亦可發現於NK細胞。
CD2分子是細胞間粘附分子,其配體分子稱為白細胞功能相關抗原-3(LFA-3,CD58),為55-70KD糖蛋白分子。可廣泛表達於造血細胞和非造血細胞。CD2分子與羊紅細胞上LFA-3結合形成花環,稱為E-花環,可用鑒定和分離人T細胞。
CD2也是信號傳導分子,可使T細胞活化,它不依賴於TCR途徑,是T細胞活化第二途徑。特別是在胸腺內早期發育階段的胸腺細胞尚未表達TCR,此時胸腺細胞的活化與增殖可能是通過CD2分子與胸腺上皮細胞表面的LFA-3分子結合而使之活化。
(六)極遲活化分子
極遲活化分子(very late activation,VLA)或稱β1粘合素(β1integrins),本族分子具有共同的β鏈(CD29),計有3種分子即VLA-4、VLA-5和VLA-6分子。它們可表達於靜止T細胞上,但活化T細胞有僅數量增多而且對特異配體的親和力也增強。VLA分子可與細胞外基質(ECM)配體分子相結合,可為T細胞活化提供協同刺激信號。VLA-4還可使淋巴細胞與Peyer小體的高內皮微靜脈以及炎症部位的內皮細胞結合,其配體分子稱為血管細胞粘附分子-1(VCAM-1)。
(七)細胞因子受體
細胞因子受體(cytokine receptor,CKR)可表達於靜止及活化T細胞表面,靜止T細胞表面的細胞因子受體親和力弱,數量少,而活化T細胞表面CKR親和力高且數量多。
T細胞表面可有多種細胞因子受體,包括IL-2R、IL-4R、IL-6R及IL-7R等。其中IL-2R由α(P55)及β(P70)鏈組成,α鏈為低親和力,β鏈為中等親和力,而αβ異聚體分子則為高親和力受體。
(八)CD44及CD45分子
CD44分子亦稱Pgp-1細胞外基質受體Ⅲ或Hermes分子。它可表達於多種細胞,包括T細胞、胸腺細胞、B細胞、粒細胞、巨噬細胞、紅細胞、神經細胞、上皮及纖維母細胞等。CD44分子可使淋巴細胞與高內皮微靜脈(HEV)結合,移行於血管,組織和淋巴之間,與淋巴細胞再循環密切相關,可視為一種歸巢受體(homingreceptor,HR)。人記憶T細胞比未受體抗原刺激的天然T細胞可表達高水平CD44分子。
CD45分子亦稱白細胞共同抗原(leukocyte common antigen)包括一組膜糖蛋白,只表達於不成熟和成熟白細胞,可包括T和B細胞、胸腺細胞、單核-巨噬細胞以及中性粒細胞。CD45分子的異構體(isofoums)常限定在某些T細胞表面表達,故稱之為CD45R。
未受抗原刺激的天然T細胞可表達CD45RA,而記憶T細胞可表達CD45RO。另外,CD45R分子胞漿區含有內源性酪氨酸磷酸酶活性,它可能對各種活化途徑具有重要調節作用。
| 關於「醫學免疫學/T細胞主要表面分子」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |