病理生理學/內生致熱原的作用方式
| 醫學電子書 >> 《病理生理學》 >> 發熱 >> 發熱的原因和機制 >> 內生致熱原的作用方式 |
| 病理生理學 |
|
|
|
無論EP是否通過血腦屏障,它在給動物靜脈內注射後,總要經過一段潛伏期才引起發熱。因而它很可能要通過某種或多個中間環節,導致調定點上移,再通過調溫反應而引起發熱。

許多學者推測有某種或某些中樞介質(也稱中樞發熱介質)參與發熱的中樞機制。先後被研究的有單胺(去甲腎上腺素、5-羥色胺)、前列腺素E(PGE)、花生四烯酸的其它衍生物,cAMP和Na+/Ca2+比值等。而最受重視的是PGE、cAMP和Na+/Ca2+比值。
(一)前列腺素
主張PGE是EP引起發熱的主要介質的現行假說的最重要依據是:①腦內(下丘腦)或ICV注射PGE引起發熱;②LP靜脈內注射或IFn ICV注射引起發熱是時,CSF中PGE2明顯增多;③下腦組織分別與LP、IFN或TNF在體外培育時,都使PGE2合成增多;④阻斷PGE合成的藥物,對LP、IFN或TNF性發熱都能解熱。
鑒於目前仍無直接證據以示LP能從外周進入腦內,因而有的學者修改了此假說,提出PGE的釋放部位是在OVLT區孔性毛細血管外周的巨噬細胞(參閱圖4-2)。LP激活後者釋出PGE,作用於OVLT區的神經元或彌散過室管膜細胞緊密連接而作用於POAH的神經元。
此外,Dinarello等(1986)認為TNF引起的雙相熱的第一相,是TNF引起下丘腦PGE增多的效應。
但是許多資料不支持PGE作為發熱介質,其根據是:(1)PGE的兩種特異拮抗物SC19220和HR546能抑止PGE性發熱,但不能抑制LP性發熱;(2)小劑量阿司匹林在抑制LP引起的CSF PGE增多的同時,可不抑制體溫上升;(3)家兔兩側POAH摘除或損傷後,向該處或ICV注入PGE均不引起發熱,但ICV注入LP仍能引起發熱,表明不需PGE參與;(4)LP注入家兔POAH,使大部分熱敏神經元敏感性受抑制,大部分冷敏神經元的敏感性提高,但PGE注入POAH,大部分熱敏神經元不受影響,約1/2冷敏神經元也不受影響;(5)MIP-1的致熱性與PGE無關。
因此,目前還難肯定PGE是EP性發熱的主要介質。
(二)cAMP
腦內有較高cAMP,也有豐富的cAMP合成降解酶系。它又是腦內多種介質的信使和突觸傳遞的重要介質,故當PGE作為發熱介質有爭議的同時,cAMP能否作為發熱介質參與中樞機制,倍受重視。十多年前國外學者積累了一些資料,支持cAMP參與發熱中樞機制,主要是:①把二丁醯cAMP給貓、兔、大鼠腦內注射,迅速引起發熱;②家兔靜脈內注射LP引起發熱時,CSF中cAMP濃度明顯增高,而環境高溫引起的體溫升高,不伴有CSF中cAMP增多。③注射茶鹼(磷酸二酯酶抑制物)在增高腦內cAMP濃度的同時,增強LP性發熱;而注射尼克酸(磷酸二酯酶激活物)則在降低cAMP濃度的同時,使LP性發熱減弱。
至於LP如何引起腦內cAMP增多,最新研究資料表明,LP可能通過提高Na+/Ca2+比值,再引起腦內cAMP增多。
(三)Na+/Ca2+比值
實驗表明,用生理鹽水替換人工腦脊液作動物腦室灌注時,引起了貓的體溫明顯上升,而加入CaCl2則可防止體溫上升。用等滲蔗糖溶液灌注後下丘腦,體溫無變化;若加入Na+,就引起體溫上升;若加入Ca2+,則可降溫。因而提出體溫調定點受Na+/Ca2+比值所調控,強調Ca2+濃度是調定點的生理學基礎,Na+/Ca2+比值上升可致調定點上移,並確定其敏感區位於後下丘腦。
進一步實驗證明,靜脈內注射LP引起發熱時,增加灌注腦室的人工腦脊液中的Ca2+濃度,可抑制發熱效應。若把灌注液改為等滲蔗糖溶液,則靜脈內注射LP不引起發熱,表明LP可能通過提高下丘腦Na+/Ca2+比值,使調定點上移而啟動調溫反應,引起體溫上升。在應用放射性同位素鈉和鈣的實驗中發現,發熱時下丘腦組織內Na+/Ca2+比值上升。
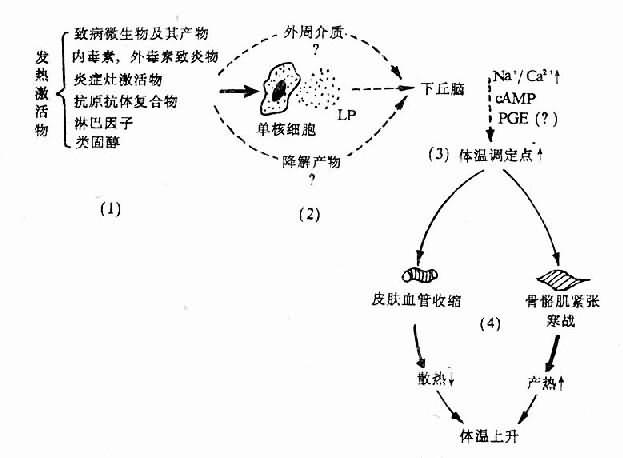
圖4-3 發熱發病學基本環節示意圖
(未包括新發現的EP)
lP如何引起Na+/Ca2+比值上升,Na+/Ca2+比值上升又如何引起調定點上移,尚缺乏深入研究。但最近國內學孝者的研究證明,用降鈣劑(EGTA)灌注側腦室引起發熱時,CSF的cAMP明顯增多;若事先灌注CaCl2,可使EGTA性體溫升高被制止,而且CSF中cAMP的增多也明顯受抑制,體溫變化與cAMP濃度變化呈明顯正相關。繼而又發現事先給家兔側腦室灌注CaCl2,不但抑制靜脈內注射LP引起的體溫上升,而且抑制了LP引起的CSF中cAMP的增多,體溫變化也與cAMP濃度變化呈明顯正相關。因此提出:《LP→下丘腦Na+/Ca2+↑→cAMP↑》可能是多種致熱原引起發熱的重要共同途徑。
總之,發熱的發生機制比較複雜,有不少細節仍未查明,但主要的或基本的環節已比較清楚。概括起來,多數發熱發病學的第一環節是激活物的作用,但至今其作用方式所知不多;第二環節,即共同的中介環節主要是EP。後者有多種,它們可能以不同給合或先後作用於POAH,或作用於外周靶細胞,再通過發熱介質參與作用;第三環節是中樞機制,無論EP是否直接進入腦內,很可能要在下丘腦通過中樞介質才引起體溫調定點上移,也不排除激活物的降解產物或外周介質到達下丘腦參與作用;第四環節是調定點上移後引起調溫效應器的反應。此時由於中心溫度低於體溫調定點的新水平,從體溫調節中樞發出調溫指令抵達產熱器官和散熱器官,一方面通過運動神經引起骨骼肌的緊張度增高或寒戰,使產熱增多;另一方面經交感神經系統引起皮膚血管收縮,使散熱減少;由於產熱大於散熱,體溫乃相應上升直至與調定點新高度相適應。這些基本環節可用下列模式圖加以表示(圖4-3)。
| 關於「病理生理學/內生致熱原的作用方式」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |