生理學/骨骼肌收縮的外部表現和力學分析
| 醫學電子書 >> 《生理學》 >> 細胞的基本功能 >> 肌細胞的收縮功能 >> 骨骼肌收縮的外部表現和力學分析 |
| 生理學 |
|
|
|
骨骼肌在體內的功能,就是它們在受刺激時能產生縮短或(和)張力,藉以完成軀體的運動或(和)抵抗外力的作用。當肌肉克服某一外力而縮短,或肌肉因縮短而牽動某一負荷時,肌肉就完成了一定量的機械功,其數值等於它所克服的阻力(或負荷)和肌肉縮短長度的乘積;如以縮短速度乘以負荷,則得出肌肉的輸出功率。但肌肉在收縮時究竟以產生張力為主或縮短為主,以及收縮時能作多少功,則要看肌肉收縮時所遇到的負荷條件和肌肉本身的功能狀態。
肌肉在體內或實驗條件下可能遇到的負荷主要有兩種:一種是在肌肉收縮前就加在肌肉上的,如把一條肌肉順著它的肌原纖維的走行方向懸掛起來而把上端固定,再在另一端懸掛一定數量的重物,後者就是前負荷。前負荷使肌肉在收縮前就處於某種程度的被拉長狀態,使它具有一定的長度,這稱為初長度;這樣由於前負荷的不同,同一肌肉就要在不同的初長度條件下進行收縮。另一種負荷稱為後負荷,它是在肌肉開始收縮時才能遇到的負荷或阻力,它不增加肌肉的初長度,但能阻礙收縮時肌肉的縮短。可以理解,對於某一具體的肌肉來說,實驗中所加負荷、特別是前負荷不應當過大,因為後者在肌肉收縮前就可能因過度的牽拉而損傷肌肉本身的結構;至於後負荷,它在大到一定程度就足以抵抗肌肉收縮所產生的最大張力,因而肌肉不再表現縮短,出現等長收縮,亦即這時肌肉雖進行了收縮,並未有長度改變;在這種情況下繼續增加後負荷,顯然不會對肌肉的收縮有什麼影響。
據上述,能影響肌肉收縮時作功能力或其力學表現的因素至少有三個,即前負荷、後負荷和肌肉本身的功能狀態(即肌肉收縮能力)。要分析某一因素影響的最簡單辦法,就是使其他因素保持在某一恆定值而改變要觀察因素的值,得到一組數據並作成一條座標曲線來進行分析。
目錄 |
(一)前負荷或肌肉初長度對肌肉收縮的影響棗長度-張力曲線
為了保持在實驗過程中肌肉本身的功能狀態基本保持不變,通常選用代謝速度較慢的兩棲類如蛙腓腸或縫匠進行實驗,實驗布置如圖2-24A所示。肌肉在下方被固定,並且邊了一個靈敏的張力換能器來記錄肌肉收縮前和收縮後的張力產生情況;肌肉的上方連一個可移動的按鈕,可以上下移動而改變肌肉的初長度,但不論初長度固定在什麼長度,同旋鈕相連的固定桿是不能動的,這就意味著把後負荷固定在無限大時的位置,肌肉在收縮時不可能縮短而只能產生張力(即前面所說的等長收縮),於是就可以觀察初長度不同時對同一肌肉所能產生張力的影響了。
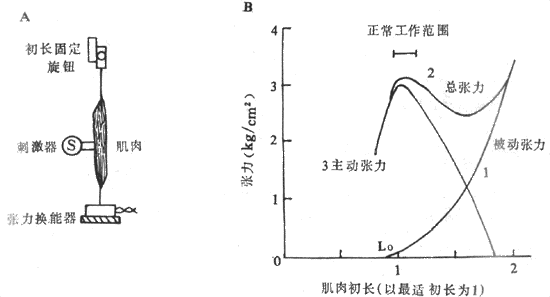
圖2-24 肌肉初長度對肌肉收縮的影響
A:在實驗布置中,下方是張力換能器,它位置固定,能把所受張力轉變為相應的
電信號,上方旋鈕可將肌肉初長在給肌肉刺激前固定於預定長度
B:3條曲線分別代表肌肉在初長度改變時的張力改變情況,被動張力指改變初長而尚未收縮的
肌肉的張力改變,總張力是在已有被動張力的基礎上肌肉收縮時產生的主動張力與前者之和
圖2-24B的長度-張力曲線反映了在依次改變肌肉的初長度時(橫座標)在張力換能器上記錄到的肌肉的張力產生的情況(縱座標)。曲線1是只改變肌肉初長度並不刺激肌肉收縮時肌肉所受的拉力,稱為被動張力曲線,它反映安靜肌肉具有某種彈性,在受到牽拉時產生某種回彈力,但牽拉超過某種程度,達到了彈性限度,被動張力急速增大,有可能造成組織損壞,其過程有如拉長一個彈簧時類似。曲線2是肌肉在具有不同前負荷即已具有被動張力的條件下進行一次收縮時記錄到的張力變化,曲線的每一點都代表那個初長度時肌肉已有的被動張力和收縮時新產生的張力之和,故整個曲線稱為總張力曲線;因此,由曲線2代表的不同初長度時的總張力減去同一初長度時的被動張力,就能得到曲線3,它表示肌肉在不同前負荷時進行收縮所能產生的張力,故稱為主動張力曲線,它反映了本實驗中要觀察的內容,即不同前負荷或初長度對肌肉收縮所能產生的張力影響:當前負荷開始增加時,每次收縮所產生的主動張力也相應地增大,但在超過某一限度後,再增加前負荷反而使主動張力越來越小,以致於為零,如曲線3右端所示。這個結論也可以表達為,對於肌肉在等長度收縮條件下所產生的主動張力大小,存在著一個最適前負荷和與之相對應的最適初長度,相當於圖2-24B橫座標上Lo的位置,在這樣的初長度情況下進行收縮,產生的張力最大。
肌肉在最適初長度條件下進行收縮何以能產生最大的張力,很容易根據肌肉被前負荷拉長時對每一肌小節中粗、細肌絲相互關係的改變來解釋。已知,肌肉產生張力和縮短,靠的是粗肌絲表面的橫橋和細肌絲之間的相互作用;肌肉初長度的大小,決定著每個肌小節的長度,亦即細肌絲和粗肌絲重疊的程度,而後者又決定於肌肉收縮時有多少橫橋可以與附近的細肌絲相互作用。從理論上分析,粗肌絲的長度是1.5μm,但在M線兩側各為0.1μm的範圍內正常時沒有橫橋,因此在M線兩側有橫橋的粗肌絲長度各為0.65μm,這樣當每側細肌絲伸入暗帶0.65μm(尚未於明帶的細肌絲長度為0.35μm),亦即肌小節總長度為2.2μm時,粗肌絲上的每個橫橋都能與細肌絲作用,因而收縮時能出現最佳的效果。當肌肉處於最適前負荷或最適初長度時,每個肌小節的長度正是2.2μm,如圖2-25箭頭3所示。如果稍稍減少前負荷使肌小節長度2.0μm(箭頭2),儘管每側細肌絲又多伸入暗帶0.1μm(這時兩側細肌絲正好相遇),但這一段正是粗肌絲上無橫橋伸出的部分,因而肌肉收縮時起作用的橫橋數目並未增多(相當於圖2-25中的箭頭2)。至於再減小肌小節的長度,則細肌絲可能穿過M線或兩側肌絲相互重合和卷屈,因而造成收縮張力下降(圖中箭頭1)。反之,如果前負荷超過最適前負荷,收縮前肌小節的長度將大於2.2μm,細肌絲和粗肌絲相互重合的程度逐漸變小,使得肌肉收縮時起作用的橫橋數也減少,造成所產生張力的下降;當前負荷使肌小節長度增加到3.5μm時,細肌絲將全部由暗帶拉出,這時肌肉受刺激時不再產生主動張力(圖中箭頭4)。由此可見,通過前負荷對肌小節中粗、細肌絲重合程度的影響,可以說明骨骼肌長度-張力曲線的特點。
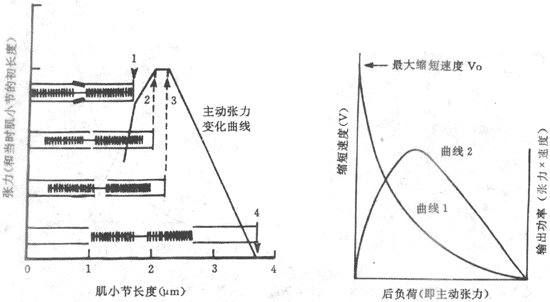
圖2-25 不同初長度時粗、細肌絲重合程度和產生張力的關係示意圖
用肌小節在不同前負荷時粗、細肌絲相對位置的改變,來說明不同前負荷時所產生的主動張力的不同:在箭頭1所指的初長度時,每個肌小節中兩側細肌絲伸入暗帶過多,互相重疊或發生卷屈,不利於與橫橋間的相互作用;在箭頭2和3所指的情況下,肌小節中全部橫橋都可與細肌絲相互作用,產生出最大主動張力;在箭頭4的情況下,細肌絲相互全部由暗帶被拉出,失去產生張力的條件
圖2-26 肌肉的張力-速度關係曲線
在肌肉前負荷固定在適當值的條件下,改變後負荷對肌肉產生張力(橫座標)
和速短速度(左側縱座標)相互關係的影響,這稱為張力-速度關係曲線(曲線1),
由此曲線可以算出不同後負荷時的輸出功率(右側縱座標),組成了曲線2
(二)肌肉後負荷對肌肉收縮的影響-張力-速度曲線
據前述,如在實驗室裝置的設計中使一條骨骼肌的前負荷固定不變而可以人為地改變後負荷,即可觀察不同後負荷對肌肉收縮的影響。一般情況下,可以把肌肉的前負荷固定在它的最適前負荷(這時出現的被動張力極小(見圖2-24B),然後,在逐次改變後負荷的情況下觀察肌肉收縮時的情況。不論在任何前負荷的情況下,如果所加後負荷超過了肌肉收縮時所能產生的最大張力(注意負荷的重量值和肌肉產生的張力的值可以用相同的物理單位度量,而且有相同的值),那麼肌肉收縮時將只產生張力而不出現肌肉長度的改變。因此,在改變後負荷的實驗中所加的後負荷都應小於這個最大張力,那麼肌肉在收縮時產生的主動張力超過這個後負荷的值時,它將會出現一定程度的長度縮短,使移動酌相同的距離,並且由於後者也可以算出一個縮短速度來(可以是初速度或平均速度)。後負荷愈小,肌肉產生的張力將較早地超過這個負荷,並且出現較大的縮短長度和縮短速度,但相當於負荷值的肌肉張力卻在縮短的過程中保持不變。這樣就得到了改變後負荷時,肌肉產生張力和其縮短速度變化的關係曲線(為了計算輸出功率,一般只分析縮短速度和張力的關係),即圖2-26曲線1,稱為張力-速度曲線。該曲線類似一條雙曲線,橫座標表示肌肉所產生的張力,縱座標表示收縮速度,雙曲線的性質則說明這二者大致呈反比的關係,即後負荷減小時,使肌肉產生的張力減小,但可得到一個較大的縮短速度;在曲線同縱軸相交的點,說明後負荷理論上為零時,可以得到該肌肉在當時的功能狀態下的最大收縮速度,在圖2-26中用V0表示;但這時因無負荷,肌肉並未作功,亦無功率輸出。在曲線同橫軸相交的點,後負荷的值相當於肌肉所能產生的最大張力,這時不能移動負荷,也沒有作功和功率輸出;在這兩個極端之間,在不同的後負荷時都能看到肌肉在產生與負荷相同的張力的情況下使負荷移動一定距離,這種類型的收縮,稱為待張收縮,都可作功和有功率輸出,但以後負荷相當於最大張力的30%左右時,肌肉的輸出功率最大,如圖2-26中曲線2所示。
(三)肌肉收縮能力的改變對肌肉收縮的影響
上述的前、後負荷的改變對肌肉收縮時張力產生、縮短速度以及作功能力等力學表現的影響,顯然是在肌肉功能狀態恆定的情況下對所處負荷條件改變所作的不同反應。但肌肉的狀態也是可以改變的,它也可以影響肌肉收縮的效率。例如,缺氧、酸中毒、肌肉中能源物質缺乏,以及其他原因引起的興奮-收縮耦聯、肌肉蛋白質或橫橋功能特性的改變,都可能降低肌肉收縮的效果,而鈣離子、咖啡因、腎上腺素等體液因素則可能通過影響肌肉的收縮機制而提高肌肉的收縮效果。將影響肌肉收縮效果的肌肉內部功能狀態的改變,定義為肌肉收縮能力(contractility)的改變,以區別於肌肉收縮時外部條件即前、後負荷改變所導致的收縮效果的改變。這樣的區分雖然在概念上比較容易,但在具體情況下要區分哪些改變是由於肌肉收縮能力的改變所引起,哪些是由於負荷條件的改變所引起,常常十分困難。例如,一個肌肉的最大張力變大了,可能是由於肌肉收縮能力的提高,但也可能是由於在這次收縮前它處於最適初長度;一個肌肉等張收縮時的收縮速度增大了,可能是由於後負荷的減小,也可能是它處於最適初長,但也可能是由於肌肉收縮能力的提高,或三者兼而有之。這就是說,很難簡單地根據肌肉某項力學指標的改變,確定是否發生了肌肉收縮能力的改變。從理論上講,肌肉收縮能力的改變對肌肉收縮的各力學表現的影響是「非選擇性」的,即收縮能力的提高可使圖2-24B中的長度-張力曲線(主動張力)和圖2-26中的張力-速度曲線的位置都升高。顯然,為了檢查收縮能力是否改變而再繪製一條條座標曲線是十分複雜的;為了簡便,如果能讓同一肌肉所處的前、後負荷條件不變而發現有肌肉收縮速度的改變,或使肌肉維持最適初長度而有最大張力的改變,則都表示肌肉收縮能力發生了改變;因為在這些條件下可以肯定,這些收縮效果的改變並不是由於前、後負荷的改變所引起,因而只能是由於肌肉的內在性能的改變引起的。
(四)肌肉的單收縮和單收縮的複合
整塊骨骼肌或單個肌細胞受到一次短促的刺激時,先是產生一次動作電位,緊接著出現一次機械收縮,後者稱為單收縮;根據收縮時肌肉所處的負荷條件不同,單收縮可以是等長的,也可以是等張的。前面敘述的肌肉收縮時各種力學表現,就是以單收縮為觀察對象而進行分析的。但在正常體內,當骨骼肌在運動神經的支配下進行自然收縮時,幾乎是無例外地接受來自神經的連續刺激,因此有必要進一步分析肌肉有受到不同頻率的連續刺激時可能發生的情況。為了便於分析,先觀察一下肌肉單收縮時電變化和機械變化在時間上的關係。圖2-27是貓脛前有一次等長單收縮時張力變化的全過程,同時記錄了肌肉的動作電位。注意圖中電反應的開始要較張力增加的開始為早,而且電變化在張力達到頂點以前早已結束;以張力最高點為界,收縮全過程可分為收縮期和舒張期,前者持續時間較後者為短。整個單收縮的時間因肌肉不同而有顯著差異,如人的眼外肌的一次單收縮不超過10ms,而腓腸肌可達100ms以上。
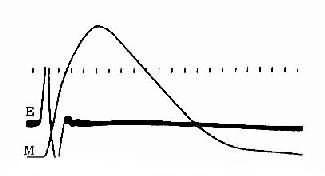
圖2-27 貓脛前肌的等長單收縮曲線
M:肌肉收縮時的張力變化曲線
E:肌肉的雙相動作電位記錄時標每 格相當於0.01s
如果給肌肉以連續的脈刺激,肌肉的收縮情況將隨刺激的頻率而有不同。如圖2-28所示,在刺激的頻率較低時,因每一個的刺激到來時由前一次刺激引起的單收縮過程(包括舒張期)已經結束,於是每次刺激都引起一次獨立的單收縮;當刺激頻率增加到某一限度時,後來的刺激有可能在前一次收縮的舒張期結束前即到達肌肉,於是肌肉在自身尚處於一定程度的縮短或張力存在的基礎上進行新的收縮,發生了所謂收縮過程的複合,這樣連續進行下去,肌肉就表現為不完全強直收縮,其特點是每次的收縮都出現在前次收縮的舒張期過程中,在描記曲線上形成鋸齒形;如果刺激頻率繼續增加,那麼肌肉就有可能在前一次收縮的收縮期結束以前或在收縮期的頂點開始新的收縮,於是各次收縮的張力或長度變化可以融合而疊加起來,使描記曲線上的鋸齒形消失,這就是完全強直收縮。
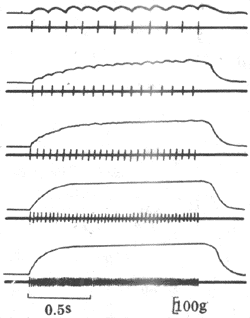
圖2-28 不同頻率的連續刺激對骨骼肌收縮的影響
每組曲線中,上方曲線是肌肉等長收縮的記錄,下方曲線是肌肉動作電位的記錄,
注意由上而下當刺激頻率依次增加時,機械收縮可出現逐漸融合,
表現不完全和完全強直收縮,但動作電位始終彼此分離,不發生融合和疊加
由於正常體內由運動神經傳到骨骼肌的興奮衝動都是快速連續的,體內骨骼肌收縮幾乎都屬於完全強直收縮,只不過強直收縮的持續時間可長可短。強直收縮顯然可以產生更大的收縮效果,例如,強直收縮所能產生的最大張力可達單收縮的4倍左右。這是因為肌肉在只接受一次刺激時,釋放到肌漿中的Ca2+很快被肌漿網上的Ca2+泵回收入肌漿網,而連續刺激可使肌漿中的Ca2+維持在一個飽和的高濃度水平。不同肌肉單收縮的持續時間不同,因而能引起肌肉出現完全強直收縮的最低臨界頻率在不同肌肉也不同,例如,單收縮快速的眼球內直肌需要每秒約350次的高頻刺激才能產生完全強直收縮,而收縮緩慢的比目魚肌只需每秒約30次的頻率就夠了。但不論在不完全強直收縮或完全強直收縮,伴隨每次刺激出現的肌肉動作電位只出現頻率加快,卻始終各自分離而不會發生融合或總和;這是由於肌肉的動作電位只持續1~2ms,當刺激頻率加速到下一次刺激落於前一次刺激引進起的動作電位持續期間時,組織又正好處於興奮的絕對不應期,這時新的刺激將無效,既不能引起新的動作電位產生,也不引起新的收縮。
| 關於「生理學/骨骼肌收縮的外部表現和力學分析」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |