生物化學與分子生物學/乳糖操縱元的表達調控
| 醫學電子書 >> 《生物化學與分子生物學》 >> 基因表達調控 >> 原核基因表達調控 >> 乳糖操縱元的表達調控 |
| 生物化學與分子生物學 |
|
|
|
如上所述乳糖操縱元的結構及其基因表達調控可綜合於圖19-7。
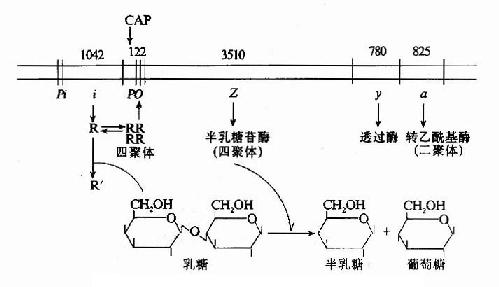
圖19-7 乳糖操縱元的結構及調控示意圖
(一)阻遏蛋白的負性調控
當大腸桿菌在沒有乳糖的環境中生存時,1ac操縱元處於阻遏狀態。i基因在其自身的啟動子Pi控制下,低水平、組成性表達產生阻遏蛋白R,每個細胞中僅維持約10個分子的阻遏蛋白。R以四聚體形式與操縱子o結合,阻礙了RNA聚合酶與啟動子P1ac的結合,阻止了基因的轉錄起動。R的阻遏作用不是絕對的,R與o偶爾解離,使細胞中還有極低水平的β-半乳糖苷酶及透過酶的生成。
當有乳糖存在時,乳糖受β-半乳糖苷酶的催化轉變為別乳糖,與R結合,使R構象變化,R四聚體解聚成單體,失去與o的親和力,與o解離,基因轉錄開放,β-半乳糖苷酶在細胞內的含量可增加1000倍。這就是乳糖對1ac操縱元的誘導作用。
一些化學合成的乳糖類似物,不受β-半乳糖苷酶的催化分解,卻也能與R特異性結合,使R構象變化,誘導1ac操縱元的開放。例如異丙基硫代半乳糖苷(isopropylthiogalactoside,IPTG)就是很強的誘導劑,不被細胞代謝而十分穩定。X-gal(5-溴-4-氯-3-吲哚-β-半乳糖苷)也是一種人工化學合成的半乳糖苷,可被β-半乳糖苷酶水解產生蘭色化合物,因此可以用作β-半乳糖苷酶活性的指示劑。IPTG和Xgal都被廣泛應用在分子生物學和基因工程的工作中。
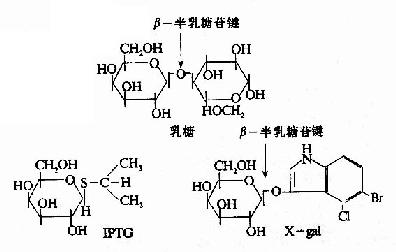
圖19-8 乳糖,IPTG和Xgal的結構
(二)CAP的正性調控
細菌中的cAMP含量與葡萄糖的分解代謝有關,當細菌利用葡萄糖分解供給能量時,cAMP生成少而分解多,cAMP含量低;相反,當環境中無葡萄糖可供利用時,cAMP含量就升高。細菌中有一種能與cAMP特異結合的cAMP受體蛋白CRP(cAMp receptor protein),當CRP未與cAMP結合時它是沒有活性的,當cAMP濃度升高時,CRP與cAMP結合併發生空間構象的變化而活化,稱為CAP(CRPcAMp activated protein),能以二聚體的方式與特定的DNA序列結合。
在1ac操縱元的啟動子P1ac上游端有一段與Plac部分重疊的序列,能與CAP特異結合,稱為CAP結合位點(CApbinding site)。CAP與這段序列結合時,可增強RNA聚合酶的轉錄活性,使轉錄提高50倍。相反,當有葡萄糖可供分解利用時,cAMP濃度降低,CRP不能被活化,1ac操縱元的結構基因表達下降。
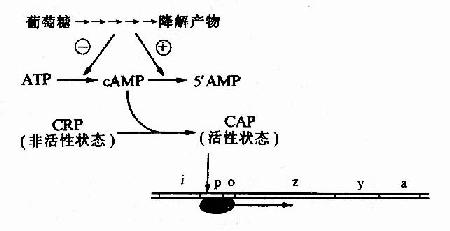
圖19-9 葡萄糖利用對乳糖操縱元的影響
由於P1ac是弱啟動子,單純因乳糖的存在發生去阻遏使1ac操縱元轉錄開放,還不能使細胞很好利用乳糖,必須同時有CAP來加強轉錄活性,細菌才能合成足夠的酶來利用乳糖。1ac操縱元的強誘導既需要有乳糖的存在,又需要沒有葡萄糖可供利用。通過這種機制,細菌優先利用環境中的葡萄糖,只有無葡萄糖而又有乳糖時,細菌才去充分利用乳糖。
細菌對葡萄糖以外的其他糖(如阿拉伯糖、半乳糖、麥芽糖等)的利用上也有類似對乳糖利的情況,在含有編碼利用阿拉伯糖的酶類基因群的阿拉伯糖操縱元(araoperon)、半乳糖操縱元(gal operon)中也有CAP結合位點,CAP也起類似的正性調控作用。所以CAP的通用名稱是分解代謝基因激活蛋白(catabolic gene activator protein)。
不難看出:CAP結合位點就是一種起正性調控作用的操縱子,CAP則是對轉錄起正性作用的控蛋白棗激活蛋白,編碼CRP的基因也是一個調控基因,不過它並不在1ac操縱元的附近,CAP可以對幾個操縱元都起作用。
從上所述,乳糖操縱元屬於可誘導操縱元(inducible operon),這類操縱元通常是關閉的,當受效應物作用後誘導開放轉錄。這類操縱元使細菌能適應環境的變化,最有效地利用環境能提供的能源底物。
| 關於「生物化學與分子生物學/乳糖操縱元的表達調控」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |