生理學/自律細胞的跨膜電位及其形成機制
| 醫學電子書 >> 《生理學》 >> 血液循環 >> 心肌的生物電現象和生理特徵 >> 心肌細胞的生物電現象 >> 自律細胞的跨膜電位及其形成機制 |
| 生理學 |
|
|
|
在沒有外來刺激時,工作細胞不能產生動作電位,在外來刺激作用下,產生一次動作電位,但兩次動作電位之間膜電位是穩定不變的。而在自律細胞,當動作電位3期復極未期達到最大值(稱最大復極電位)之後,4期的膜電位並不穩定於這一水平,而是立即開始自動除極,除極達閾電位後引起興奮,出現另一個動作電位。這種現象,周而復始,動作電位就不斷地產生。出現於4期的這種自動除極過程,具有隨時間而遞增的特點,其除極速度遠較0期除極緩慢;不同類型的自律細胞4期除極速度參差不一,但同類自律細胞4期除極速度比較恆定。這種4期自動除極(亦稱4期緩慢除極或緩慢舒張期除極),是自律細胞產生自動節律性興奮的基礎。
根據細胞膜除極的跨膜電流的基本規律可分析自律細胞4期自動除極形成的機制。不難推測,自律細胞由於淨外向電流使膜復極(3期)達最大復極電位後,在4期中又出現一種逐漸增強的淨內向電流,從而使膜內正電位逐漸增加,膜便逐漸除極。這種進行性淨內向電流的產生,有以下三種可能的原因:①內向電流的逐漸增強;②外向電流的逐漸衰退;③兩者兼有。不同類型的自律細胞,4期自動除極都是由這種進行性淨內向電流所引起,但構成淨內向電流的離子流的方向和離子本質並不完全相同。
1.浦肯野細胞浦肯野細胞是一種快反應自律細胞。作為一種快反應型細胞,它的動作電位的形態與心室肌細胞相似,產生的離子基礎也基本相同。
關於浦肯野細胞4期自動除極形成的機制,80年代研究資料表明,在浦肯野細胞,隨著復極的進行,導致膜復極的外向K+電流逐漸衰減,而同時在膜電位4期可記錄到一種隨時間推移而逐漸增強的內向電流(If)(圖4-7)。If通道在動作電位3期復極電位達-60mV左右開始被激活開放,其激活程度隨著復極的進行、膜內負電性的增加而增加,至-100mV左右就充分激活。因此,內向電流表現出時間依從性增強,膜的除極程度因而也隨時間而增加,一旦達到閾電位水平,便又產生另一次動作電位,與此同時,這種內向電流在膜除極達-50mV左右因通道失活而中止。可見,動作電位的復極期膜電位本身是引起這種內向電流啟動和發展的因素,內向電流的產生和增強導致膜的進行性除極,而膜的除極一方面引起另一次動作電位,一方面又反過來中止這種內向電流。這一連串的過程是自律細胞「自我」啟動、「自我」發展,又「自我」限制的,由此可以理解為什麼自律細胞能夠自動地、不斷地產生節律性興奮。
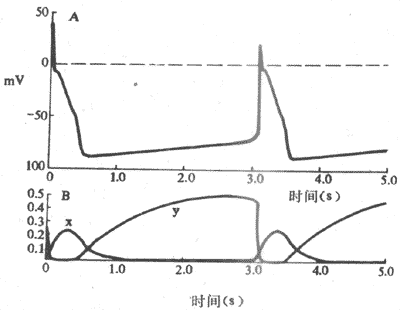
圖4-7 浦肯野細胞起搏機制
A;跨膜電位 B:由х閘門控制的Ik衰減以及由у閘門控制的If,兩者在形成起搏電位中的相對關係
這種4期內向電流,通常稱為起搏電流,其主要離子成分為Na+ ,但也有K+參與。由於使它充分激活的膜電位為-100mV,因而認為,構成起搏內向電流的是一種被膜的超極化激活的非特異性內向(主要是是Na+)離子流,標誌符號為If。If的通道允許Na+通過,但不同於快Na+通道,兩者激活的電壓水平不同;If可被銫(Cs)所阻斷,而河豚毒卻不能阻斷它。目前,關於If及其通道的研究資料尚有若干不能充分予以解釋的疑點,對If的進一步研究正受到心肌電生理學者們的高度關注。
2.竇房結細胞的跨膜電位及其形成機制 竇房結含有豐富的自律細胞,動作電位復極後出現明顯的4期自動除極,但它是一種慢反應自律細胞,其跨膜電位具有許多不同於心室肌快反應細胞和浦肯野快反應自律細胞的特徵:①竇房結細胞的最大復極電位(-70mV)和閾電位(-40mV)均高於(電位較正)浦肯野細胞;②0期除極結束時,膜內電位為0mV左右,不出現明顯的極化倒轉;③其除極幅度(70mV)小於浦肯野細胞(為120mV),而0期除極時程(7ms左右)卻又比後者(1-2ms)長得多。原因是竇房結細胞0期除極速度(約10V/s)明顯慢於浦肯野細胞(200-1000V/s),因此,動作電位升支遠不如後者那麼陡峭;④沒有明顯的復極1期和平台期;⑤4期自動除極速度(約0.1V/s)卻比浦肯野細胞(約0.02V/s)要快,記錄曲線上竇房結細胞4期膜電位變化的斜率大於浦肯野細胞.圖4-8顯示心室肌快反應細胞與竇房結細胞跨膜電位變化的差別。
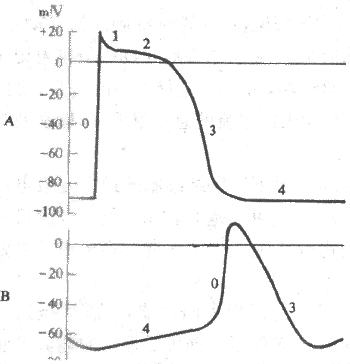
圖4-8 心室肌(A)與竇房結(B)細胞跨膜電位的比較
竇房結細胞的直徑很小,進行電生理研究有一定困難。直到70年代中期,才開始在竇房結小標本上採用電壓鉗技術對其跨膜離子流進行了定量研究,但目前尚未能充分闡明它的跨膜電位,尤其是4期起搏電流的離子基礎。學者們觀察到,竇房結細胞0期除極不受細胞外Na+濃度的影響,對河豚毒很不敏感;相反,它受細胞外Ca2+濃度的明顯影響,並可被抑制鈣通道的藥物和離子(如異搏定、D-600和Mn2+等)所阻斷。據此可以認為,引起竇房結細胞動作電位0期除極的內向電流是由Ca2+負載的。這種內向電流被稱為第二內向電流;而引起快反應細胞(心室肌、心房肌和浦肯野細胞)0期除極的快Na+內流稱為第一內向電流。根據已有的研究資料,可將竇房結細胞動作電位的形成過程描述如下:當膜電位由最大復極電位自動除極達閾電位水平時,激活膜上鈣通道,引起Ca2+內向流(Ica),導致0期除極;隨後,鈣通道逐漸失活,Ca2+內流相應減少;另一方面,在復極初期,有一種K+通道被激活,出現K+外向流(Ik)。Ca2+內流的逐漸減少和K+外流的逐漸增加,膜便逐漸復極。由「慢」通道所控制、由Ca2+內流所引起的緩慢0期除極,是竇房結細胞動作電位的主要特徵,因此,相應稱為慢反應細胞和慢反應電位,以區別於前述心室肌等快反應細胞和快反應電位。
竇房結細胞的4期自動除極也由隨時間而增長的淨內向電流所引起,但其構成成分比較複雜,是幾種跨膜離子流的混合。目前已知,在竇房結細胞4期可以記錄到三種膜電流,包括一種外電流和兩種內向電流,不過它們在竇房結細胞起搏活動中所起作用的大小以及起作用的時間有所不同。
(1)Ik通道的激活和逐漸增強所造成的K+外向流,是導致竇房結細胞復極的原因。Ik通道在膜復極達-40mV時便開始逐漸失活,K+外流因此漸漸減少,導致膜內正電荷逐漸增加而形成4期除極。目前認為,由於Ik通道的時間依從性逐漸失活所造成的K+外流進行性衰減,是竇房結細胞4期自動除極的最重要的離子基礎(圖4-9);
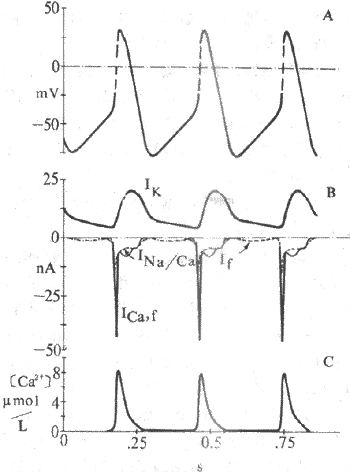
圖4-9 竇房結動作電位和起搏電位的離子機制
A.跨膜電位 B.越膜電位C. 胞漿Ca2+濃度表示動作電位升支由Ica,
f構成,起搏電位 由Ik和If及Ina/Ca構成
(2)If:If是一種進行性增強的內向離子(主要為Na+)流,在浦肯野細胞起搏活動中,If起著極重要的作用,而Ik衰減的作用很小。與此恰相反,竇房結細胞4期雖也可記錄到If ,但它對起搏活動所起的作用不如Ik衰減。實驗證明,用Cs2+選擇性阻斷If後,竇房結自發放頻率僅輕度減少;對家兔竇房結細胞4期淨內向電流的總幅值而言,Ik衰減與If兩者所起作用的比例為6:1。
If通道的最大激活電位為-100mV左右,而正常情況下竇房結細胞的最大復極電位為-70mV,在這種電位水平下,If通道的激活十分緩慢,這可能是If在竇房結4期除極過程中所起作用不大的原因。若竇房結細胞發生超級化時,If則可能成為起搏電流中的主要成分。
(3)此外,竇房結細胞4期中還存在一種非特異性的緩慢內向電流,在膜除極達-60mV時被激活,可見,它在自動除極過程的後1/3期間才起作用。這種緩慢內向電流可能是生電性Na+-Ca2+交換的結果(Na+-Ca2+交換時,心肌細胞排出一個Ca2+,攝入3個Na+,出/入細胞正電荷之比為2:3,形成內向電流)。
心肌細胞的類型 除了按照功能和電生理特性將心肌細胞分為工作細胞和自律細胞之外,還可以根據其生物活動的特徵,特別是動作電位0期除極的速度,將心肌細胞分為快反應細胞和慢反應細胞,其動作電位相應稱為快反應電位和慢反應電位;然後再結合其自律性,可將心肌細胞分為以下四種類型:
快反應非自律細胞:包括心房肌細胞和心室肌細胞;
快反應自律細胞:浦肯野自律細胞;
慢反應自律細胞:竇房結自律細胞,以及房結區和結希區的自律細胞;
慢反應非自律細胞:結區細胞。
心肌細胞的跨膜離子流 由多種離子通過細胞膜上的特異性或非特異性離子通道以及經離子交換轉運形成的跨膜離子運動,是心肌細胞跨膜電位形成的基礎。心肌細胞跨膜離子流的種類眾多,性質很複雜,近年又取得許多進展。由於在實驗研究中對這些離子通道的命名和認識比較混亂,甚至前後矛盾,故將目前比較肯定的主要幾種離子流和通道的名稱、存在的部位和作用歸納介紹如下:
1.內向離子流
(1)INa:稱為快速Na+流,存在於快反應細胞,是引起快反應細胞0期除極的內向電流(稱第一內向電流)的離子基礎。
(2)Isi:稱為緩慢內向電流,也稱第二內向電流,過去認為它是一種由慢通道控制的緩慢Ca2+流,故標誌為ICa,其功能是構成快反應細胞的平台期和慢反應細胞的除極期。新近的研究結果在很大程度上修正了對Ica的認識。目前認為,第二內向電流並不是單一的Ca2+流,而是由特性各異的三個組分所構成;第一組分稱Ica.f,二、三組分別稱Is1.2和Ica.s。其中①Ica.f是一種快速Ca2+流,其通道激活和失活的速度遠比已往所認為的要快得多。它融合於快反應細胞Na+內流的最後部分,共同形成動作電位升支的上段,而對平台期的作用很小。它是慢反應細胞除極的離子基礎。對工作細胞而言,肌漿網Ca2+再生性釋放帥細胞外Ca2+進入胞漿觸發的,既然Ica.f的激活十分迅速,那麼Ca2+釋放的觸發有及興奮-收縮耦聯的啟動速度都比以往認為的要快得多;②第三組分Ica.s,是一種較Ica.f微弱而緩慢的Ca2+流,主要作用是維持快反應細胞平台期;③Is1.2稱第二內向電流第二組分,其離子本質不甚清楚,可能是Na+-Ca2+交換的生電電流,故也稱為INa.Ca,在平台期起作用;慢反應自律細胞4期自動除極晚期也有它的作用。
(3)If:超極化激活的非特異性內向離子流,主要由Na+攜帶,存在於自律細胞4期。If是決定浦肯野快反應自律起搏活動的主要負離子流,而在竇房結慢反應自律細胞起搏活動中,其作用不如Ik衰減。
2.外向離子流
(1)Ikl:存在於快反應細胞,是決定快反應工作細胞靜息電位的離子流,並在復極2期和3期起複極作用;
(2)Ik:這種外向電流主要由K+攜帶,但也有Na2+參加,不是單純的K+流,故又稱Ix,在快反應細胞復極3期起重要作用,故又稱為復極電流,因而也決定著浦肯野細胞的最大復極電位;Ix也存在於慢反應自律細胞,促使膜復極,4期內呈進行性衰減,是形成4期除極的主要離子基礎;
(3)Ito:是快反應細胞1期復極的離子流,離子成分主要為K+,也有Na+參與。
3.化學門控離子通道 近年相繼證實,在心肌細胞膜上,除了電壓門控通道外,還存在化學門控離子通道;它們的作用並不在於維持和產生正常跨膜電位(靜息電位和動作電位),但對於心肌細胞活動的調節以及異常電活動的電生產有著特別重要的意義。重要的化學門控離子通道有以下三種;
(1)乙醯膽鹼控制的K+通道(Ik-ACh):早年曾認為ACh激活的是Ikl,近年發現是通過G蛋白激活開放了另一種與在生物物理學特性和生理學特性上均不相同的K+通道。
(2)ATP依從性K+通道(Ik-ATP):ATP的作用並不是分解供能激活此通道,而是維持此通道在正常情況下處於關閉狀態。當心肌細胞內ATP降到臨界水平以下時(如心肌缺血時),此種特殊的K+通道開放。大量K+外漏以致缺血心肌細胞局部高鉀而引起除極,誘發心律失常。硫脲類藥物可阻斷此通道。
(3)INa–k—Ca2+通道:是細胞內Ca2+增高時激活的一種非特異性正離子通道,載流離子是Na+和K+,形成一過性內向離子流(Iti)。實驗表明,在某些情況下,浦肯野細胞在動作電位復極後可產生一種除極電位(延遲後除極電位),當它達閾電位時就可以誘發另一個新的動作電位,形成異位搏動。Iti就是延遲後除極電位的離子基礎。洋地黃中毒,細胞外低K+或低Na+,以及咖啡因、兒茶酚胺等可引起細胞內Ca2+超負荷的因素,均可誘發或加強Iti和延遲後除極電位。
| 關於「生理學/自律細胞的跨膜電位及其形成機制」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |