核酸熱力學
| A+醫學百科 >> 核酸熱力學 |
核酸熱力學是指溫度影響雙鏈DNA(dsDNA)的核酸結構。DNA變性(DNA denaturation)又稱DNA融化(DNA melting)是DNA雙螺旋解開成為兩條單股長鏈的過程。在過程中,使兩股長鏈上的鹼基相連的氫鍵會斷裂。 DNA的變性可以是溫度升高而產生的作用,也可能是其他化學物質如尿素的誘導。使DNA解開的融化溫度(Tm)是依DNA鏈的長度,以及特定核苷酸序列的組成形式而定。
目錄 |
基礎概念
雜交
雜交是建立在兩個以上序列具有特異性相互作用的互補鏈之間產生非共價鍵結核酸成單一複合物,此種情況稱為DNA雙股螺旋。寡核苷酸,DNA或RNA將在正常條件下綁定到其互補之序列,而使此互補鏈可以容易地與彼此結合。為了降低差異和獲得能量最高的優選配合物,實驗室中使用一種叫做退火的方法。然而,由於不同的核甘酸分子具有不同的幾何形狀,兩條鏈之間的差異不一會使它們之間具有較低的能量親和性來進行結合。通過量化兩條鏈在退火之間的相似性可以提供鹼基序列中溫度測量不相容效果的信息。該複合物可以通過熱解離來進行變性,亦被稱為融化,此法為將複合物溶液加熱,打破其核酸鹼基對間的氫鍵,而使兩股分開。在沒有外來的不利因素,雜交和融化的過程可以重複連續下去,其中配對方法可參考聚合酶鏈式反應。最常見的是,核酸鹼基對以A = T和G≡C的方式成對形成,其中後者是較穩定的(由於氫鍵數較多)。
變性
DNA變性,又稱DNA解鏈或融化,是由雙鏈去氧核糖核酸開旋並通過斷裂分離成單鏈疏水且堆疊的單鏈分子。「變性」還可以指DNA鏈的誘導化學物質,如分離尿素。
DNA變性的方法亦可用於分析DNA的某些方面。因為胞嘧啶/鳥嘌呤鹼基配對通常比腺嘌呤/胸腺嘧啶鹼基配對更強,胞嘧啶和鳥嘌呤在一個基因組中(稱為「GC含量 「),可以通過測量在該基因組DNA的熔解溫度來估計。[1]較高的溫度與GC含量高有關。由於GC含有三個氫鍵,而AT只具有兩個氫鍵。
DNA變性,也可用於檢測兩個不同的DNA序列之間之序列差異。將DNA加熱和變性成單鏈狀態,並將該混合物冷卻使可以重新進行雜交。雜交分子的相似序列中如果互補序列有差異,則會導致鹼基配對中斷。在基因組範圍中,該方法已被用於估算兩物種之間遺傳距離的研究,稱為DNA-DNA雜交。[2] 在其中的單個分區的DNA,變性梯度凝膠和溫度梯度凝膠可用於檢測此兩個序列,此法稱為溫度梯度凝膠電泳,為表現較小差異時使用的方法。[3][4]
DNA熔解的也應用於分子生物學技術,特別是在聚合酶鏈式反應。儘管此技術不能診斷DNA熔化的溫度,所以估計(Tm)是確定來調整是和溫度是非常重要的。DNA的熔解溫度也可被用作用於均衡的一組分子的雜交優勢,例如的寡核苷酸探針DNA微陣列。
退火
退火,在遺傳學中,用於單鏈的DNA或RNA的一互補序列由氫鍵形成配對的雙鏈多核苷酸。該術語通常用來描述一個的結合的DNA探針,或一個的結合的引子對DNA鏈中一個聚合酶鏈式反應。這個詞也經常說是蛋白質,如RAD52可以幫助DNA退火。
兩種形態的熱力學
以下幾個公式用於計算(Tm)的值。[5][6]某些公式更精確地預測DNA雙鏈體的解鏈溫度。[7]對於DNA寡核苷酸,兩個型態的進過程短的DNA序列能被準確地雜交熱力學描述,假設兩個單鏈中間互相彼此結合成雙鏈的機率為100%之下,可以極簡單地描述熱力學單鏈核酸A和B形成雙鏈核酸AB
- AB ↔ A + B
該反應的平衡常數為![K=\frac{[A][B]}{[AB]}](/images/math/b/7/e/b7e00ed422db77d8353553c23cf2601a.png) ,根據范特霍夫等式,其自由能 ΔG, 和 K 是 ΔG° = -RTln K,其中R是理想氣體常數,T是該反應的開爾文溫度。此法適用於核酸系統。
,根據范特霍夫等式,其自由能 ΔG, 和 K 是 ΔG° = -RTln K,其中R是理想氣體常數,T是該反應的開爾文溫度。此法適用於核酸系統。
![\Delta G^\circ = -RT\ln\frac{[A][B]}{[AB]}](/images/math/b/5/b/b5bdf3ee29d5fd2d7faf9d2acd790a61.png) .
.
於溫度達到(Tm)一半時解鏈就已經開始發生。如果沒有額外的核酸存在時,[A],[B]和[AB]將是相等的,並且等於雙鏈核酸的初始濃度的一半。這給出了一個核酸雙鏈體的融點的表達式:
![T_m = -\frac{\Delta G^\circ}{R\ln\frac{[AB]_{initial}}{2}}](/images/math/6/d/6/6d6af6e77e8e3639dba2a4b083a02d62.png) .
.
由於 ΔG° = ΔH° -TΔS°, Tm 也可以以下式表示:
![T_m = \frac{\Delta H^\circ}{\Delta S^\circ-R\ln\frac{[AB]_{initial}}{2}}](/images/math/e/3/d/e3dbd30c248ab9cf12043d0916819dca.png) .
.
ΔH° and ΔS°通常為合作而非分解反應,則演變成下式:[8]
![T_m = \frac{\Delta H^\circ}{\Delta S^\circ+R\ln([A]_{total} - [B]_{total}/2)}](/images/math/1/c/2/1c2a52bccb83259d3db0d3253088274a.png) , where [B]total < [A]total.
, where [B]total < [A]total.
如上述所提到的,該方程是基於只有雙鏈和隨機捲曲兩種狀態參與了熔化的假設。然而,核酸還可以通過許多中間狀態的融化。考慮到這類複雜的行為,必須結合統計力學,特別是長序列相關的複合物。
此外還有 Wallace method For 15-20 nucleotides primers
Tm = 2(A+T) + 4(G+C) ℃
從核酸序列推定熱力學性質
上段表示了熔化溫度和熱力學參數都是相互關聯的(ΔG° or ΔH° & ΔS°)。從觀察熔化溫度中可以通過實驗確定其熱力學參數。當一個給定的核酸序列的熱力學參數是,熔融溫度即可以預測。對寡核苷酸而言,這些參數可以很好地通過最近鄰模型近似。
最近鄰近似法(Nearest-neighbor method)
在不同鏈之之間的相互作用一定程度上取決於相鄰鹼基。最近鄰模型將DNA螺旋表示為「鄰近」的鹼基對之間的相互作用的序列組。[8] 例如,下面所表示的DNA具有最接近由箭頭表示鄰相互作用。
- ↓ ↓ ↓ ↓ ↓
- 5' C-G-T-T-G-A 3'
- 3' G-C-A-A-C-T 5'
此DNA來自其自身之自由能ΔG於37℃時表示為
ΔG°37(理想) = ΔG°37(CG 最初) + ΔG°37(CG/GC) + ΔG°37(GT/CA) + ΔG°37(TT/AA) + ΔG°37(TG/AC) + ΔG°37(GA/CT) + ΔG°37(AT 最初)
第一項代表第一鹼基對,CG的自由能,第二項包括形成第二個鹼基對,GC的雙方的自由能,以此類推。該鹼基對產生與先前的鹼基對之間的堆積作用。在一般情況下,形成的核酸雙鏈體的自由能總和以下式表示:
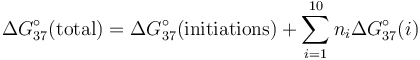 .
.
上式亦可表達成以下形式:
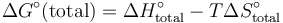 .
.
ΔH° 和 ΔS°的值已经确定至少有十个可能的相互作用。(表一)
| 最近鄰序列 (5'-3'/3'-5') |
ΔH° 千焦/莫耳 |
ΔS° 焦耳/(莫耳·K) |
ΔG°37 千焦/莫耳 |
|---|---|---|---|
| AA/TT | −33.1 | −92.9 | −4.26 |
| AT/TA | −30.1 | −85.4 | −3.67 |
| TA/AT | −30.1 | −89.1 | −2.50 |
| CA/GT | −35.6 | −95.0 | −6.12 |
| GT/CA | −35.1 | −93.7 | −6.09 |
| CT/GA | −32.6 | −87.9 | −5.40 |
| GA/CT | −34.3 | −92.9 | −5.51 |
| CG/GC | −44.4 | −113.8 | −9.07 |
| GC/CG | −41.0 | −102.1 | −9.36 |
| GG/CC | −33.5 | −83.3 | −7.66 |
| 終端AT鹼基對 | 9.6 | 17.2 | 4.31 |
| 終端的GC鹼基對 | 0.4 | −11.7 | 4.05 |
表1所示的十個鄰居相關的參數是由短寡核苷酸的雙鏈熔解點確定的。特別的是,他們十組之中只有八組是獨立的。 最近鄰模型可以擴展超出了華森克立克的配對,包括錯配和鄰近鹼基對之間相互作用的參數。[9]這使得含有孤立錯配的序列具有熱力學參數的估計,例如像下列序列(箭頭表示不匹配):
- ↓ ↓ ↓
- 5' G-G-A-C-T-G-A-C-G 3'
- 3' C-C-T-G-G-C-T-G-C 5'
參考文獻
- ↑ M. Mandel and J. Marmur. Use of Ultraviolet Absorbance-Temperature Profile for Determining the Guanine plus Cytosine Content of DNA. Methods in Enzymology. Methods in Enzymology. 1968, 12 (2): 198–206. doi:10.1016/0076-6879(67)12133-2. ISBN 978-0-12-181856-2.
- ↑ C.G. Sibley and J.E. Ahlquist. The Phylogeny of the Hominoid Primates, as Indicated by DNA-DNA Hybridization. Journal of Molecular Evolution. 1984, 20 (1): 2–15. doi:10.1007/BF02101980. PMID 6429338.
- ↑ R.M. Myers, T. Maniatis, and L.S. Lerman. Detection and Localization of Single Base Changes by Denaturing Gradient Gel Electrophoresis. Methods in Enzymology. Methods in Enzymology. 1987, 155: 501–527. doi:10.1016/0076-6879(87)55033-9. ISBN 978-0-12-182056-5. PMID 3431470.
- ↑ T. Po, G. Steger, V. Rosenbaum, J. Kaper, and D. Riesner. [http//www.ncbi.nlm.nih.gov/pmc/articles/PMC305948/ Double-stranded cucumovirus associated RNA 5: experimental analysis of necrogenic and non-necrogenic variants by temperature-gradient gel electrophoresis]. Nucleic Acids Research. 1987, 15 (13): 5069–5083. doi:10.1093/nar/15.13.5069. PMID 3601667. PMC 305948.
- ↑ Breslauer, K.J. et al.; Frank, R; Blöcker, H; Marky, LA. [http//www.ncbi.nlm.nih.gov/pmc/articles/PMC323600/ Predicting DNA Duplex Stability from the Base Sequence]. Proc. Natl. Acad. Sci. USA.. 1986, 83 (11): 3746–3750. doi:10.1073/pnas.83.11.3746. PMID 3459152. PMC 323600. (pdf)
- ↑ Rychlik, W.; Spencer, W. J.; Rhoads, R. E.. [http//www.ncbi.nlm.nih.gov/pmc/articles/PMC332522/ Optimization of the annealing temperature for DNA amplification in vitro]. Nucleic Acids Res.. 1990, 18 (21): 6409–6412. doi:10.1093/nar/18.21.6409. PMID 2243783. PMC 332522.
- ↑ Owczarzy R., Vallone P.M., Gallo F.J., Paner T.M., Lane M.J. and Benight A.S. Predicting sequence-dependent melting stability of short duplex DNA oligomers. Biopolymers. 1997, 44 (3): 217–239. doi:10.1002/(SICI)1097-0282(1997)44:3<217::AID-BIP3>3.0.CO;2-Y. PMID 9591477. (pdf)
- ↑ 8.0 8.1 8.2 John SantaLucia Jr.. [http//www.ncbi.nlm.nih.gov/pmc/articles/PMC19045/ A unified view of polymer, dumbbell, and oligonucleotide DNA nearest-neighbor thermodynamics]. Proc. Natl. Acad. Sci. USA. 1998, 95 (4): 1460–5. doi:10.1073/pnas.95.4.1460. PMID 9465037. PMC 19045.
- ↑ John SantaLucia Jr., John; Donald Hicks. The thermodynamics of DNA structural motifs. Annual Review of Biophysics and Biomolecular Structure. June 2004, 33: 415–440 [27 March 2013]. doi:10.1146/annurev.biophys.32.110601.141800. PMID 15139820.
參見
外部連結
|
|||||||||||
參考來源
| 關於「核酸熱力學」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |