生理學/興奮在同一細胞上的傳導機制
| 醫學電子書 >> 《生理學》 >> 細胞的基本功能 >> 細胞的興奮性和生物電現象 >> 興奮的引起和興奮的傳導機制 >> 興奮在同一細胞上的傳導機制 |
| 生理學 |
|
|
|
可興奮細胞的特徵之一是它任何一處的膜產生的動作電位,都可沿著細胞膜向周圍傳播,使整個細胞的膜都經歷一次類似於被刺激部位的離子電導的改變,表現為動作電位沿整個細胞膜的傳導。傳導的機制實際已包含在興奮膜的上述特性之中。設想一條槍烏賊的無髓神經纖維的某一小段,因受到足夠強的外加刺激而出現了動作電位(圖2-18,B左端),即該處出現了膜兩側電位的暫時性倒轉,由靜息時的內負外正變為內正外負,但和該段神經相鄰接的神經段仍處於安靜時的極化狀態;由於膜兩側的溶液都是導電的,於是在已興奮的神經段和與它相鄰的未興奮的神經段之間,將由於電位差的存在而有電荷移動,稱為局部電流。它的運動方向是:膜外有正電荷由未興奮段移向已興奮段,膜內有正電荷由已興奮段移向未興奮段。這樣流動的結果,是造成未興奮段膜內電位升高而膜外電位降低,亦即引起該處膜的去極化;這一過程開始時,就相當於電緊張性擴布。根據上述關於興奮產生的機制的分析,當任何原因使膜的去極化達到閾電位的水平時,都會大量激活該處的Na+通道而導致動作電位的出現。因此,當局部電流的出現使鄰接的未興奮的膜去極化到閾電位時,也會使該段出現它自己的動作電位。所謂動作電位的傳導,實際是已興奮的膜部分通過局部電流「刺激」了未興奮的膜部分,使之出現動作電位;這樣的過程在膜表面連續進行下去,就表現為興奮在整個細胞的傳導。由於鋒電位產生期間電位變化的幅度和陡度相當大,因此在單一細胞局部電流的強度超過了引起鄰近膜興奮所必需的閾強度數倍以上,因而以局部電流為基礎的傳導過程是相當「安全」的,亦即一般不易因某處動作電位不足以使鄰接的膜產生興奮而導致傳導「阻滯」,這一點與一般化學性突觸處的興奮傳遞有明顯的差別。
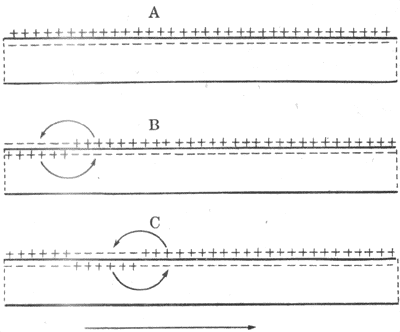
圖2-18 神經纖維傳導機制的模式圖彎箭頭表示膜內外
局部電流的流動方向,下方直箭頭表示衝動傳導方向。
A:靜息時 B:發生興奮後 C:傳導過程中
興奮傳導機制雖然以無髓神經纖維為例,但在其他可興奮細胞(如骨骼肌細胞)的興奮傳導,基本上遵循同樣的機制。有髓神經纖維在軸突外麵包有一層相當厚的髓鞘,髓鞘主要成分的脂質是不導電或不允許帶電離子通過的,因此只有在髓鞘暫時中斷的朗飛結處,軸突膜才能和細胞外液接觸,使跨膜離子移動得以進行。因此,當有髓纖維受到外加刺激時,動作電位只能在鄰近刺激點的朗飛結處產生,而局部電流也只能發生在相鄰的朗飛結之間,其外電路要通過髓鞘外面的組織間液,因此,動作電位表現為跨過每一段髓鞘而在相鄰朗飛結處相繼出現,這稱為興奮的跳躍式傳導。
跳躍式傳導時的興奮傳導速度,顯然比上述無髓纖維或一般細胞的傳導速度快得多;而且由於跳躍式傳導時,單位長度內每傳導一次興奮所涉及的跨膜離子運動的總數要少得多,因此它還是一種「節能」的傳導方式。看來,神經髓鞘的出現是進化過程中既能增加神經纖維傳導速度、又能減少生物能量消耗的一種方式。無脊椎動物沒有有髓神經纖維,而無髓纖維增加增加傳導速度的一個可能途徑是增大軸突的直徑,因為這樣可以減少膜內液體的電阻而增加局部電流的強度,使動作電位的傳導速度加快;這大概就是需要進行快速神經反應的槍烏賊在進化中出現巨大的無髓神經纖維的道理所在。但徐科(1993)等人指出,某些無脊椎動物的神經纖維也可以一種特殊的方式進行跳躍式傳導。
如果一條神經纖維在它的中間部受到刺激,將會有動作電位由中間向纖維兩端傳送,這是由於局部電流可以出現在原興奮段兩側之故。由此可以理解,興奮在同一細胞上的傳導,並不限於朝向某一方向;體內神經纖維所以有傳入和傳出之分,只是由於在整體的自然條件下,傳入纖維只能在它們和感受器相連接的外周端被刺激,而傳出纖維只能在它們的細胞體產生衝動而傳向外周,並非是由於這些纖維本身只能單方向傳導興奮的緣故。以動作電位為興奮出現的指標,可以測定興奮在各種細胞的傳導速度。例如,人體一些較粗的有髓神經纖維的傳導速度,最快可達每秒m以上,而一些細胞的無髓纖維每秒傳導距離還不到1m;構成心臟內部傳導系統的浦肯野細胞,每秒傳導速度約4~5m,是心肌細胞中傳導速度最快的。
| 關於「生理學/興奮在同一細胞上的傳導機制」的留言: | |
|
目前暫無留言 | |
| 添加留言 | |